
威远江自然保护区思茅松种群的林窗更新研究
2013-06-23 13:56:28廖迎芸彭明春党承林
环境科学导刊 2013年2期
廖迎芸,彭明春,党承林
(1.云南省环境科学研究院,云南昆明650034;2.云南大学,云南昆明650091)
威远江自然保护区思茅松种群的林窗更新研究
廖迎芸1,彭明春2,党承林2
(1.云南省环境科学研究院,云南昆明650034;2.云南大学,云南昆明650091)
在野外考察的基础上,运用径级结构、高度结构的方法对思茅松种群的林窗更新进行研究,研究结果表明:思茅松纯林的林窗更新与灌草层盖度,尤其是草本层盖度密切相关。灌木层盖度的大小影响幼苗的存活,灌木层盖度小,幼苗存活率高。草本层盖度的大小影响种子的萌发,草本层盖度小,种子萌发率高。在针阔混交林林窗中,当常绿阔叶树稀少或土层浅薄时,思茅松更新个体年龄差异大,当常绿阔叶树长势好且密度较大时,思茅松更新个体年龄普遍差异较小。
思茅松;种群;林窗;更新
思茅松是云南的 “特有植物”,经济价值高,是不可取代的遗传材料[1]。云南省虽然建立了威远江自然保护区保护思茅松种源,但至今保护区内开展的科学研究不多[2]。本次研究通过对思茅松种群的林窗更新研究,为思茅松种质资源的保护和森林资源有效管理提供科学依据。
1 研究地概况
威远江自然保护区位于云南省景谷县境内。保护区地理位置为北纬23。06′~23。17′,东经100。31′~100。35′,距景谷县城61km。保护区属无量山脉威远江水系,境内地形分割破碎起伏较大,山高坡陡,为典型的中心切割窄谷地形。海拔多在1000~1500m,最高点1734m。保护区内的土壤中赤红壤占最大比例。由于高温多雨,水热同季,有机质分解较快,土壤腐殖质层较薄,自然肥力较低。根据景谷县气象站资料,当地年均气温20.1℃,极端最高气温38.6℃,极端最低气温0.2℃,年总降水量1026~1402mm,年蒸发量1601~2022mm。
保护区内及周边主要为思茅松纯林和针阔混交林,季风常绿阔叶林有少量分布。其中分布面积最大的是思茅松纯林,群落结构分乔木、灌木、草本三层。乔木层以思茅松(Pinus kesiya var.langbianensis)为优势种,其他物种数量少,偶见有黄毛青冈(Cyclobalanopsis delavayi)、滇青冈(C.glau-coides)、毛叶青冈(C.kerrii)、栓皮栎(Quercus variabilis)、槲栎(Q.aliena)。灌木层常见种类有:母猪果(Helicia nilagirica)、臭荚蒾(Viburnum foetidum)、余甘子(Phyllanthus emblica)、米饭花(Vaccinium sprengelii)、大叶算盘子(Glochidion lanceolarium)、盐肤木(Rhus chinensis)、野漆(Toxicodendron succedaneum)、水锦树(Wendlandia parviflora)、云南黄杞(Engelhardtia spicata)、卵叶南烛(Lyonia ovalifolia)、滇银柴(Aporusa yunnanensis)、黑面神(Breynia fruticosa)等。草本层常见种类有:三点金草(Desmodium triflorum)、长尖莎草(Cyperus cuspidatus)、斑茅(Saccharum arundinaceum)、无芒竹叶草(Oplismenus compositus)、紫茎泽兰(Eupatorium adenphorum)、宿包豆(Shuteria involucrata)、拟鳞毛蕨(Kuniwatsukia cuspidata)。
针阔混交林以思茅松、黄毛青冈、华南石栎(Lithocarpus fenestratus)构成,分布面积小,结构分乔木上层、乔木下层、灌木层、草本层四层。组成物种数相对丰富,常见种类有毛果柃木(Eurya trichocarpa)、高山栲(Castanopsis delavayi)、栓皮栎、西南桦(Betula alnoides)、滇银柴、米饭花、盐肤木、水锦树、野牡丹(Melastoma candidum)、余甘子、西南杭子梢(Campylotropis delavayi)、酸藤子(Embelia laeta)、红梗润楠(Machilus rufipes)、茶梨(Anneslea fragrans)、云南黄杞、卵叶南烛、灰毛浆果楝(Cipadessa cinerascens)、思茅蒲桃(Syzygium szemaoense)、白牛胆(Inula cap-pa)、小槐花(Catenaria caudata)、耳草(Hedyotis auricularia)、金发草(Pogonatherum paniceum)、毛姜花(Hedychium villosum)、沿阶草(Ophiopogon bodinieri)、长尖莎草、臭黄皮(Clausena emarginata)、紫茎泽兰、硬秆子草(Capillipedium assimile)、大叶千斤拔(Flemingia macrophylla)、棕叶芦(Thysanolaena maxima)、鸢尾(Iris tectorum)。
2 研究内容和方法
2.1 研究内容
广义上的更新即演替,涉及一个较大的时间尺度,存在较多的争议。因此,本次研究针对狭义的更新,即对思茅松种群 (思茅松纯林、思茅松针阔混交林)进行更新研究。
2.2 研究方法
2.2.1 野外样地调查
个体年龄:采取人工伐倒思茅松树后数树干基部的年轮来确定。
林窗年龄:由于林窗形成木高度腐烂,难以从腐烂程度来确定其真实年龄,因此林窗年龄采用选取林窗内较大的思茅松2~3棵,计算其个体年龄平均值来估算。
林窗面积:选取面积相近的不同林窗,从林窗中心向外划定一块10×10m即面积为100m2的区域作为林窗更新的样地代表以减少分析中的变量。
(1)思茅松纯林的林窗更新
根据调查选取了3个不同灌草层盖度的林窗样地Q1~Q3。分别记录3个样地的环境概况、群落特征以及样地中每株思茅松的胸径、高度、冠幅、生活力 (分强、中、弱3个等级)。
(2)思茅松针阔混交林的林窗更新
根据调查选取了2个针阔混交林林窗样地Q4~Q5,分别记录2个样地的环境概况、群落特征以及样地中每株思茅松的胸径、高度、冠幅、生活力。
生活力又称生活强度或茂盛度,主要反映生态上的适应和竞争能力,不包括因物候原因而生活力变化者。一般分为3级:
生活力强:植株高度较高,枝叶繁茂,生长旺盛,在群落中生长势很好;
生活力中:植株高度中等,一般低于最高植株0.5m~1.0m,生长势一般;
生活力弱:植株高度较矮,一般低于生活力中等植株0.5m~1.0m,枝叶稀疏,生长柔弱。
2.2.2 数据处理
(1)径级结构图的编制
径级结构对于认识更新机制非常重要且效果良好[3~4],本研究采用径级结构代替年龄结构,将调查的胸径(DBH)以2cm为一个径级划分标准作出径级结构图[5]。即0cm<Ⅰ级≤2cm,2cm<Ⅱ级≤4 cm,4cm<Ⅲ级≤6cm,6cm<Ⅳ级≤8 cm,8cm<Ⅴ级≤10cm,10cm<Ⅵ级≤12 cm,12cm<Ⅶ级≤14cm,14cm<Ⅷ级≤16 cm。
(2)高度结构图的编制
高度结构的划分标准为:0m<Ⅰ级≤1.5m,1.5m<Ⅱ级≤3.5m,3.5m<Ⅲ级≤5.5m,5.5m<Ⅳ级≤7.5m,7.5m<Ⅴ级≤9.5m。
(3)环境因子与更新的关系
根据样地环境概况,比较群落小环境 (海拔、坡度、坡向等)的差异及其与更新的关系。
3 研究结果及分析
本研究共调查了5个样地,其中思茅松纯林3个,针阔混交林2个,样地概况见表1。
3.1 思茅松纯林的林窗更新
选取的3个不同灌草丛盖度样地的群落特征见表2。

表1 调查样地的环境概况
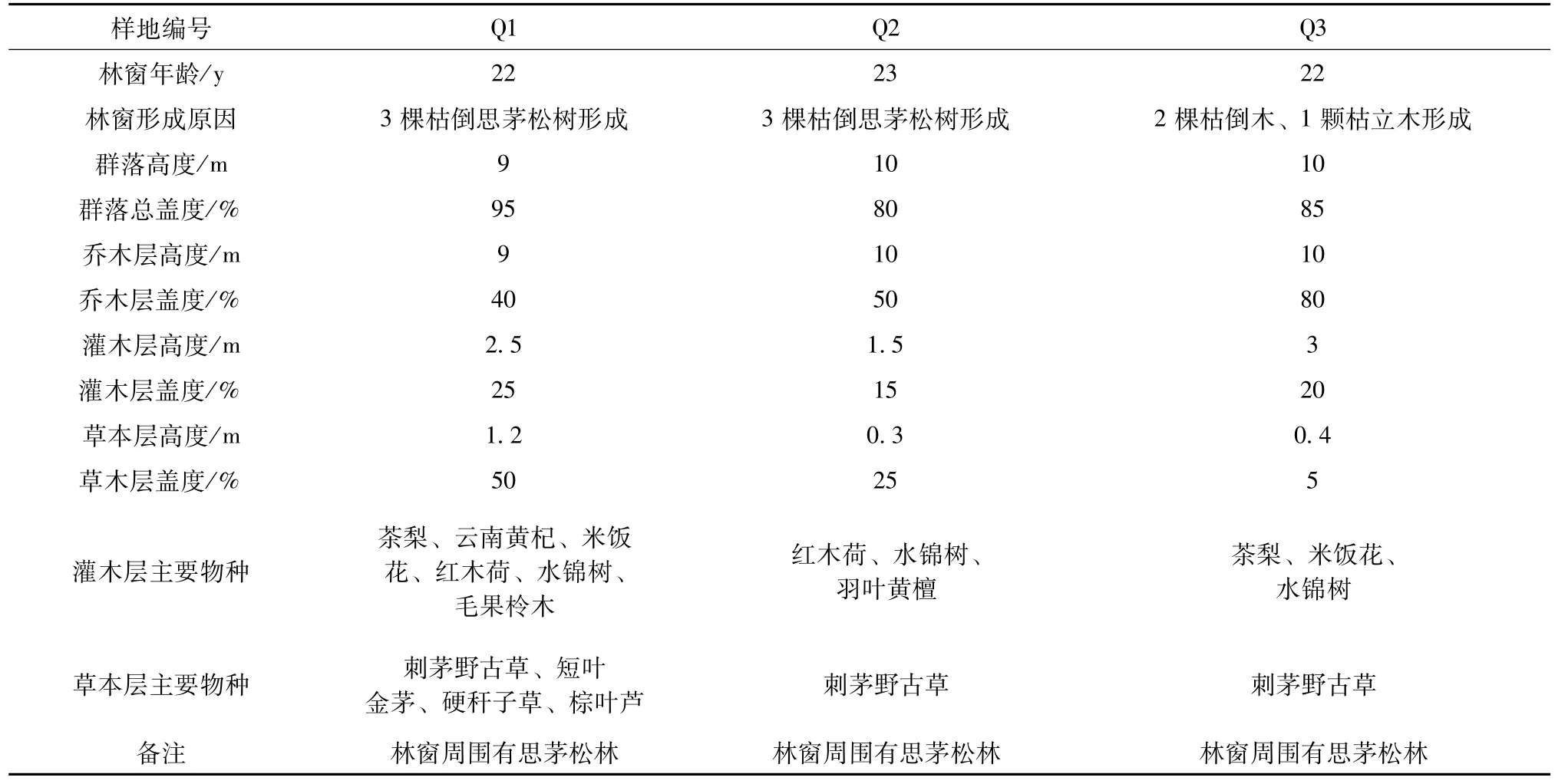
表2 不同灌草丛盖度的样地群落特征表
样地Q1(见图1)中思茅松种群径级结构为倒金字塔形,共有思茅松51株,Ⅴ级和Ⅳ的数量较多,占29.4%和27.5%;Ⅵ级的数量稍少于Ⅴ级;从Ⅴ级到Ⅰ级数量逐步递减,到Ⅰ级仅剩2株,占3.9%。形成该径级结构的原因是林窗形成后光照强度增大、光照时间增长,有利于林窗种子库中的思茅松种子萌发,位于Ⅵ级的思茅松最早萌发,由于没有上层物种的遮挡,能够迅速占领林窗空地的上层空间,但这一级定居的思茅松数量不多,随着周围思茅松林的进一步补充种子,定居的思茅松数量逐渐增加,因此,Ⅴ级和Ⅳ级的数量多于Ⅵ级。从Ⅴ级到Ⅰ级数量逐步递减的最重要原因是,林窗形成后不断增加最后高达75%灌草层盖度尤其是高达50%的草本层盖度,显著地抑制了思茅松种子的萌发、生长。而且随着早期定居的思茅松的生长、林窗的逐步郁闭,林下光照减弱,传播来的种子即使萌发也会因光照达不到光补偿点而死亡。
样地Q2(见图2)和样地Q3(见图3)的径级结构除Ⅰ级数量稍少外,整个结构为金字塔形。其中Q2内共有思茅松51株,Ⅱ级和Ⅲ级数量最多分别占39.2%和27.5%,Ⅰ级和Ⅴ级数量最少,占7.8%和5.9%。样地Q3内共有思茅松79株,Ⅰ级有10株,Ⅱ级和Ⅲ级数量最多,分别占31.7%和30.4%,往后数量减少,到Ⅶ级仅有1株,占1.3%。形成该径级结构的原因是初期传播进来的种子数量较少,且缺少适宜种子萌发生长的土壤,因此成功萌发生长的幼苗数量比较少。Ⅱ级到Ⅲ级的数量最多,说明两个种群均有较大的幼苗储备,主要原因是低盖度灌草层尤其是低密度的草本层有利于种子萌发。幼苗存活率高。这种较高的幼苗储备和存活率为思茅松在林窗内实现良好更新提供了基础。
比较三者的环境条件,海拔相差不到100m且样地Q1的海拔更接近思茅松良好更新的海拔范围。样地Q1虽处于半阴 (东北)坡上,光照没有半阳(西南)坡充足,但由于其乔木层盖度为40%,比其它两个样地低,因此3个样地林下光照差别不大。比较3个样地的灌木层盖度分别为25%、15%和20%,差异不大,主要是草本层盖度差异较大,分别为50%、25%和5%,呈现递减趋势,因此可以认为灌丛层盖度尤其是草本层盖度是造成三者径级结构差异的主要原因。灌草层对更新的影响主要有以下两个方面:
当灌草层盖度尤其是草本层盖度很高的时候,对更新有极大的影响,表现在3个方面:①灌木层盖度高说明灌木层物种生长繁茂,占据了绝大部分空间,即使思茅松种子能够成功萌发,也因缺乏适宜的生境面临难以更新的困境。②草本层盖度高意味着它们会截留每年补充的很大比例的思茅松种子,加上草本层在表土层形成极密的草根盘结以及枯枝落叶形成的地被物减少了种子接触到土壤的机率,从而降低了种子萌发的机率,而种子的萌发是实现更新的首要条件。③灌草层共同组成的林下层在某种程度上可以改变微环境,如凋落物的厚度、光照、温度以及土壤湿度等。较高的灌草层盖度产生了较多的枯枝落叶物,即使在枯枝落叶层上成功萌发的种子,在干旱季节也会由于水分供给的有限性,林窗内光照低于其光补偿点,或其它可利用资源的缺乏,导致幼苗死亡率高。
当灌草层盖度尤其是草本层盖度中等和低的时候,草本层阻隔的种子比高盖度的草本层阻隔的要少,而且部分截留的种子还可能被雨水冲刷或被风吹动而落入土壤,因此有较多的种子能够萌发。而且灌木层内物种稀疏,群落内有可利用空间,种内和种间竞争相对减弱,大大提高了更新的成功率。
可见,影响更新不同阶段的主导因子不同,其中影响种子萌发的主导因子是草本层盖度。较高的草本层盖度严重影响了种子的萌发,无法迈出种群更新的第一步。而较低的草本层盖度则有利于使较多的种子直接接触地面土壤,更新较好。灌木层盖度大小则影响幼苗的存活,较低的灌木层盖度表明还有可利用的空间和资源,有利于幼苗生长。此外,幼苗能否正常生长发育还受到群落微环境、环境容量等多个因素的影响。
3.2 针阔混交林的林窗更新
根据调查样地的径级结构和高度结构图,存在种群内个体间年龄差异大和个体间年龄差异小的不同现象。
3.2.1 林窗内思茅松年龄差异大
林窗是约18年前由3棵思茅松树自然枯倒后形成的。该林窗处于下坡位,环境异质性比较明显。林窗北向地段较陡,母岩裸露,土层浅薄贫瘠,有雨水冲刷形成的多条大小不一的小冲沟;而南向地段坡度相对平缓,地表有少量的灌木草本植物。
样地Q4为思茅松、麻栎、华南石栎的针阔混交林。群落总盖度70%,其中乔木层高9m,盖度70%。除建群种外,还有滇银柴、余甘子、米饭花;灌木层高3m,盖度15%;草本层缺乏,高0.5m,盖度仅为10%。
径级结构图中各个阶段均有分布,但位于林窗南向的9棵思茅松和其它常绿阔叶树种长势好,思茅松最高达9.0m,比其它常绿阔叶树种高1m左右,除1株思茅松仅为3.5m外其余均高于6.0m,平均胸径为8.2cm;而位于林窗北向的裸露地带上仅长有3株思茅松幼苗和1株幼树,除幼树高7.0m外,幼苗均低于3.0m,平均胸径为3.0cm。
比较径级结构图和高度结构图,可以看出位于林窗南北两侧的思茅松在高度和胸径上差异较大,位于南向最早进入林窗的思茅松和位于北向高度仅为0.3m的一年生幼苗年龄差异可达到17年左右。位于林窗南向的思茅松数量多,平均年龄大,位于林窗北向的思茅松数量少,平均年龄小,长势差。形成这种差异的原因是同一个林窗内,环境并非完全同一和均质,位于南向的思茅松在高度上普遍高于北向的思茅松,说明了两侧思茅松在该林窗内更新时间上的不等性。
导致这种更新时间上的不等性是由于以下3个原因单独或综合作用的结果:①在土壤条件相对较好和相对平缓的南向地段,进入林窗的思茅松和常绿阔叶树种的种子很快萌发长成幼苗,群落环境随之发生改变,林下逐渐阴蔽且该群落处于半阴(西北)坡上,后期进入的思茅松种子在荫蔽的环境下难以萌发或存活,因此位于南向的思茅松年龄较大且林下幼苗在林窗形成一定时间后数量极少。②在林窗北向裸露地段上,由于常绿阔叶树种的种子较大,在坡度为25°的山坡上容易受重力作用向下滚落,偶有种子能够停留下来,也因土壤浅薄贫瘠、缺少水分等恶劣环境条件而难以萌发或存活。③下落在该裸露地段上的思茅松种子暴露于地表,也相对增大了被啮齿动物采食的机率,保留下来的种子数量少。然而思茅松在长期的进化过程中,发展了自身应对不良环境条件的机制。它每年都生产大量种子,发育成熟的种子总有机会落入裸露地面的缝隙中或冲沟以外的较平缓的小斑块中,一旦条件适宜,思茅松种子就迅速萌发生长,但比之水热土壤条件好的生境,其生长速度受到一定影响。另外,这种地段有比较充裕的空间供生长,因此出现了数量少且高矮参差不齐的思茅松在裸露地段上。
3.2.2 林窗内思茅松年龄差异小
林窗是约13年前由2棵思茅松大树自然枯倒后形成的。样地Q5是以思茅松、西南桦、茶梨为共优种的针阔混交林。群落总盖度达85%,其中乔木层高10m,盖度75%,思茅松和西南桦比茶梨高2~3m,灌木层高5m,盖度20%,主要种类有水锦树、云南黄杞;草本层高1m,盖度40%,枯枝落叶层厚约2cm。
样地Q5中共有思茅松幼树12株,年龄差异最大为4年,其中生活力强的6株(包括1株11年生、4株9年生、1株8年生),平均高度8.5m,较茶梨等常绿阔叶树高1.5m;生活力中等的4株(包括1株11年生、2株8年生、1株7年生),平均高度6.7m,和常绿阔叶树相差不大;生活力差的2株(包括1株8年生、1株7年生),平均高度5.9m,较常绿阔叶树低约1m,此外还有一些阔叶树幼苗和2株胸径<2.5cm的枯死的思茅松幼苗。
样地中树种是林窗形成后侵入的,乔木层中思茅松和西南桦都是速生树种,它们均高居于常绿阔叶树种之上;而茶梨、水锦树生长速度稍慢,高度上低于思茅松和西南桦。
早进入林窗的思茅松比晚进入林窗的思茅松普遍表现出生活力强的特征。这是因为早进入林窗的思茅松比常绿阔叶树高1.5m,更容易获取光照资源;而晚进入林窗的思茅松高度上低常绿阔叶树约1m,容易受到高思茅松和阔叶树种的荫蔽,难以获得充足的光照,长势差。对于个别年龄相同但生活力不同的思茅松,如:样地中同是8年生的思茅松却表现出强、中、弱3种生活力特征,这主要是种子遗传本身的差异,其次与周围个体的竞争也有关系。遗传性状好的个体在周围物种竞争中将能够获得优势,长势良好。
林窗内无7年生以下的幼苗,其原因是随着早期思茅松和其它阔叶树种的定居生长,思茅松幼苗的生长受到强烈的抑制。当林窗郁闭后,林下光照十分微弱,仅存的2株思茅松幼苗在这种环境中难以存活,最终枯死,因而乔木层中思茅松年龄差异小,草本层中无幼苗,灌木层中则相继出现了耐荫性较强的阔叶树幼种。但是随着林窗的郁闭,思茅松种群的更新可以通过在当地的陡坡上,现有群落上层的枯倒木或风倒木产生 “多米诺效应”——第一株树撞倒第二株树上,第二株树接着又撞倒第三株树,这样一株大树树倒能产生比它本身所占空间更大的林窗来开始新的进程。
4 结语
在威远江自然保护区的管理中,对于思茅松纯林,采取天然更新为主、人工抚育间伐为辅。当种群密度过高,尤其是灌草丛盖度较高时,伐除其中矮小、弯曲、断稍、有病害的植株,避免造成种群个体小、种质资源差的局面。
对于针阔混交林,若常绿阔叶树种生长过于繁茂,应保留树桩合理伐除其中一部分生长旺盛的常绿阔叶树种,形成林窗,同时让阔叶树进行萌生。当常绿阔叶树种的萌生枝再次形成较大的荫蔽时,应再次对萌生枝进行部分修剪,这样便增大了林下的光照强度,为思茅松更新创造条件,不仅有利于思茅松种质资源的保护,而且有利于常绿阔叶树种发挥保护生物多样性和水土保持的功能。
[1]邹天才.贵州特有种子植物种质资源与利用评价研究 [J].林业科学,2001,37(5):46.
[2]吴兆录.思茅松研究现状的探讨[J].林业科学,1994,30(2):151-156.
[3]Shimano K.A power function for forest structure and regeneration pattern of pioneer and climax species in patch mosaic forests[J]. Plant Ecology,2000,(146):207.
[4]闫桂琴.秦岭太白红杉种群结构与动态的研究 [J].应用生态学报,2001,12(6):824.
[5]胡万良.人工诱导阔叶红松林种群结构及分布格局[J].东北林业大学学报,2000,28(3):56.
[6]F.H.鲍尔曼.森林生态系统的格局与过程[M].北京:科学出版社,1985:139.
A Study on the Gap Generation of Pinus kesiya var.langbianensis in Weiyuanjiang Nature Reserve
LIAO Ying-yun1,PENG Ming-chun2,DANG Cheng-lin2
(1.Yunnan Institute of Environmental Science,Kunming Yunnan 650034 China)
Based on the field survey,the gap generation of the population is analyzed through DBH structure and height structure.The results show that the Pinus kesiya var.langbianensis generation in gaps is related to the shrub and herb cover,especially the herb cover.The shrub cover influences the survive of seedlings,and the low shrub cover induces high germination of the seeds.In the gap of coniferous and broad-leaved mixed forest,the age of the pine individuals differs greatly when the evergreen broad-leaved trees are sparse and the soil is thin;on the contrary,the pine individuals differ slightly in age when the evergreen broad-leaved trees are densely and well grown.
Pinus kesiya var.langbianensis;population;gap;regeneration
X173
:A
:1673-9655(2013)02-0020-06
2012-11-20
廖迎芸 (1981-),女,汉族,工程师,主要从事生态学相关研究。
猜你喜欢
广西林业科学(2022年4期)2022-11-07 01:50:34
西南林业大学学报(2022年5期)2022-10-17 09:56:38
林业调查规划(2022年3期)2022-06-13 09:31:04
广西林业科学(2021年6期)2022-01-21 07:39:10
东北林业大学学报(2021年1期)2021-12-03 19:17:48
东北林业大学学报(2021年1期)2021-01-14 04:26:10
农业机械学报(2020年1期)2020-03-13 00:27:38
西部林业科学(2018年4期)2018-02-14 22:43:50
山东林业科技(2017年1期)2017-06-29 07:53:58
上海农业学报(2016年5期)2016-02-10 06:53:24
