
野豌豆属牧草种质花粉形态的扫描电镜观察
2013-04-25 09:30:16马利超刘志鹏
草业科学 2013年11期
刘 鹏,马利超,王 宇,刘志鹏
(兰州大学草地农业科技学院 草地农业生态系统国家重点实验室,甘肃 兰州 730020)
野豌豆属(Vicia)牧草具有较高的营养价值和较强的抗寒特性,适宜在高寒和高海拔地区栽培,是高原牧区重要的蛋白饲料来源[1]。因其具有生物产量高、生育周期短、种子粗蛋白含量高[2]和抗寒性较强等特点[3]而被科技工作者广泛关注。有报道显示,蚕豆(V.faba)是我国最早栽培的豆类作物之一,可作饲料、绿肥和蜜源植物种植,其种子蛋白质含量高达22.35%,是优质的植物粗蛋白源。山野豌豆(V.amoena)除作为优良牧草及绿肥外,其种子还有一定的药用价值。长柔毛野豌豆(V.villosa)种子中含有丰富的植物凝血素,这一物质在免疫学、肿瘤生物学、细胞生物学及发育生物学的研究中已得到了广泛的应用。箭筈豌豆(V.sativa)为一年生或越年生叶卷须半攀援性草本植物,整株可用作绿肥及饲料。对野豌豆属植物的花粉生物学特性研究,可为植物物种及其种质资源鉴定提供重要的形态依据。野豌豆属植物约有190个物种[4],Ball[5]将该属划分成4个组:Vicia,Cracca,Ervum和Faba。随后研究人员进一步分析了Vicia和Vicilla两个亚属的生物学特性[6-7]。此外,Vicia亚属包括很多具有重要农业价值的牧草[8]。Maxted[9]在原表型分类的基础上又将Vicia亚属细分为9大系列,38个物种,14个亚种。本属植物主要分布在北半球温带地区,拉丁美洲有零星分布[10]。根据中国数字植物标本馆(http://www.cvh.org.cn/cms/)和宋敏等[11]的统计结果,在我国本属植物广布于全国各省,华北、西北和西南地区较多,其中栽培种5种,野生种36种,华南和台湾地区分布很少,仅有野生种1种。
关于野豌豆属植物花粉形态的研究,国内学者曾开展过一些工作。苏亚拉图等[12]观察了我国内蒙古境内13种野豌豆属植物花粉扫描电镜的结果,发现野豌豆属植物的花粉形态、萌发孔和外壁纹饰等特征在种内是稳定的。野豌豆属新变种狭柳叶野豌豆(V.venosavar.ich-hsinganica)花粉形态为长球形,具三孔沟,沟缘加厚,孔膜高升,沟浅,外壁具不规则网状纹饰[13]。Erdtman[14]认为利用花粉粒形态特征可以提高植物分类的精确度。深入观察花粉的形态特征可为植物分类学和授粉生物学等研究提供有价值的参考信息[15-16]。虽然很多学者在野豌豆属植物花粉形态上做了一定的研究工作,但仍存在一些不足,如苏亚拉图[12-13]共观察了13种内蒙古野豌豆属及其新变种植物的花粉形态,但13个物种全部来自中国内蒙古,具有地域限制。本研究收集了中国、德国、埃塞俄比亚和蒙古的野豌豆属花粉材料,使野豌豆属孢粉学资料具有广域性,更加完善。本研究利用高分辨率扫描电镜,较为系统地对来自4个国家的8份野豌豆属植物材料的花粉形态进行观察,获取新材料的花粉形态学信息,探讨其系统分类学特性,以期为野豌豆属植物的孢粉学研究[17]积累相关信息。
1 材料和方法
1.1试验材料 野豌豆属植物种子由美国国家种质资源库(NPGS)和兰州大学草地农业科技学院提供。于2012年8月初播种于兰州大学榆中校区智能温室(16 h光照,8 h黑暗;光照时22 ℃,黑暗时20 ℃)。在当年10-11月盛花期分别采集成熟花药作为扫描电镜观察的材料。材料具体名称及来源地见表1。
1.2试验方法
1.2.1花粉采集 植物盛花期进行取样,每个物种采集5朵鲜花,用镊子将花药取出,放入1.5 mL EP管中,常温下干燥48 h后存放于4 ℃冰箱中。为了获得自然真实的表面纹饰,采用自然散出的花粉扫描,个别材料的花粉是从刺破的成熟花药中释放得到的,扫描前均未经任何试剂处理[18]。在导电胶上将花粉均匀散布,置于金属载物台上,用常规真空喷镀法喷金90 s后,置于扫描电镜(日本电子光学公司,JSM-5600LV)下观察,选取有代表性的视野,分别进行显微拍摄。
1.2.2计算方法 每份材料随机取30粒花粉进行测量和统计,取平均值表示变化幅度。花粉粒大小以极轴长乘以赤道轴长(P×E)来表示,其形状以极轴长和赤道轴长的比值(P/E)来表示。花粉的表面纹饰形态特征参照Punt等[19]的描述,P/E为0.5~0.8是扁圆形,P/E为0.8~1.0是扁球形,P/E为1.0~1.1是长球形。

表1 8份野豌豆属种质名称及其来源Table 1 Name and origin of eight Vicia accessions
2 结果
对8份野豌豆属植物材料的花粉进行扫描电镜观察发现,野豌豆属植物的花粉为单粒花粉,赤道面观是扁球形的有5种(V.amoena、V.fabavar.faba、V.fabavar.minuta、V.sativa2560A和V.sativa2560B)、扁圆形的2种(V.sativa2505和V.sativa2556)和长球形的1种(V.villosa)(表2)。对花粉粒大小的比较发现,不同物种花粉粒大小差异比较明显,V.amoena和V.sativa2556极轴长显著大于其他材料(P<0.05)。V.sativa2556赤道轴长显著大于其他材料(P<0.05)。此外,野豌豆属植物花粉外壁表面纹饰可分为网状纹饰(V.fabavar.faba、V.fabavar.minuta、V.villosa和V.sativa2505)、脑状纹饰(V.sativa2556和V.sativa2560A)、片状纹饰(V.sativa2560B)和穴状纹饰(V.amoena)4种类型,其极面观为三裂圆形。萌发孔沟接近两极且宽窄不一。
2.1萌发孔形态特征 本研究所观察的8份植物材料花粉具有下列几种萌发孔特征:1)萌发沟细窄,沟中部有明显凸起。如V.fabavar.faba(图1 A~C),V.sativa2560A(图2 D~F),V.villosa(图3 D~F),V.sativa2556(图4 A~C),V.fabavar.minuta(图4 D~F)。2)萌发沟较宽,沟中部无凸起。如V.sativa2505(图2 A~C),V.sativa2560B(图3 A~C)。3)萌发沟中部无凸起。如V.amoena(图1 D~F)。
2.2外壁表面纹饰 所观察的花粉外壁表面纹饰有以下4个类型:1)网状纹饰。网纹较深,网眼大小不一,如V.fabavar.faba(图1 A~C);网纹较浅,网眼细密,如V.fabavar.minuta(图4 D~F);网纹较浅,网眼较大,如V.villosa(图3 D~F)、V.sativa2505(图2 A~C)。2)脑状纹饰。纹饰较深,立体感强烈,如V.sativa2560A(图2 D~F);纹饰较浅,如V.sativa2556(图4 A~C)。3)穴状纹饰。如V.amoena(图1 D~F)。4)片状纹饰。如V.sativa2560B(图3 A~C)。
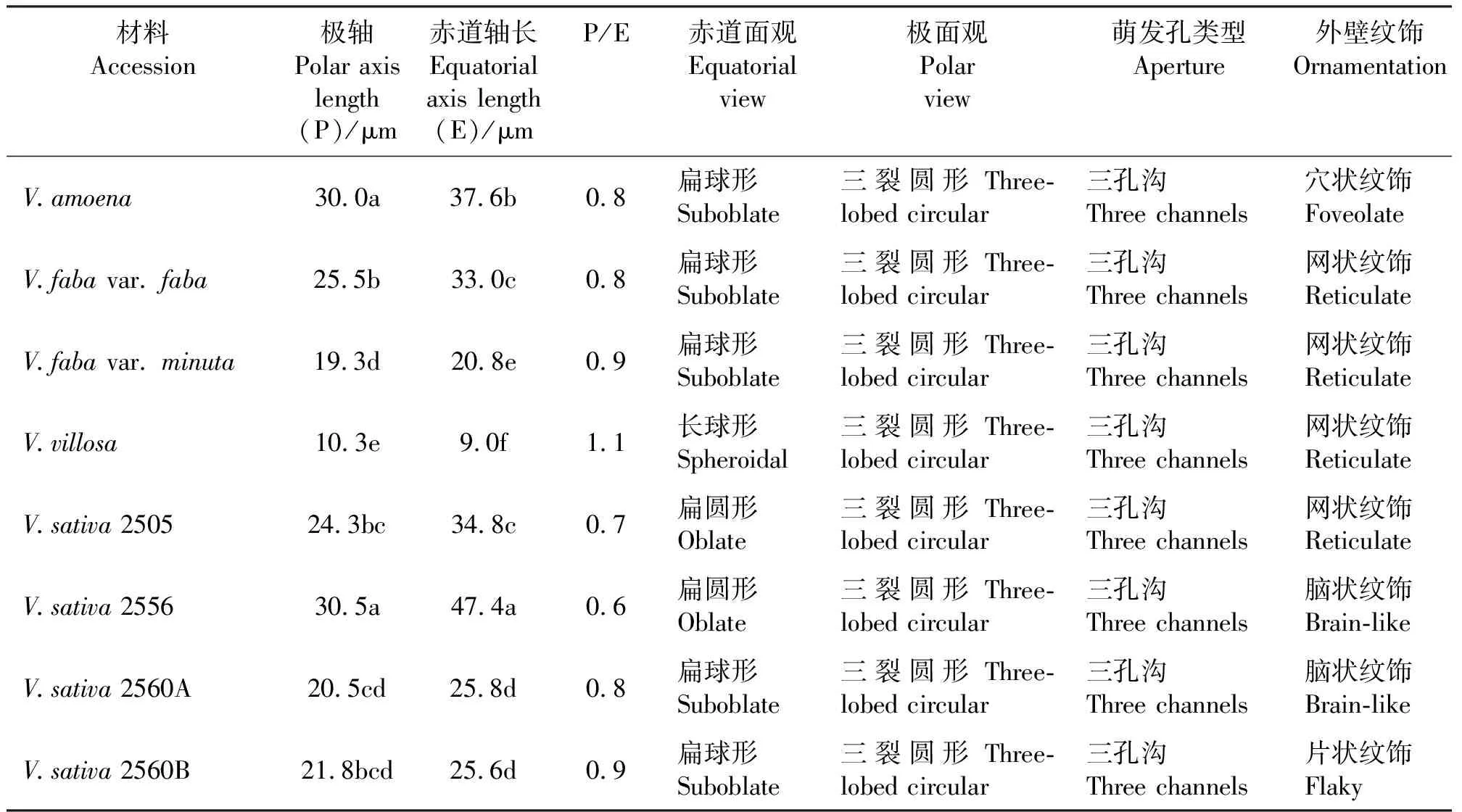
表2 8份野豌豆属植物材料花粉形态学特征Table 2 Main characteristics of the pollen morphology in eight accessions of Vicia

图1 V.faba var. faba(A~C)和V.amoena(D~F)花粉粒形态Fig.1 Pollen morphology of V.faba var. faba(A~C)and V.amoena(D~F)
V.fabavar.faba(图1 A~C)极轴长25.5,赤道轴长33.0,P/E值为0.8。赤道面观为扁球形,极面观为三裂圆形。三孔萌发沟并延伸达两极,萌发沟细窄,沟中部有明显凸起。表面为网状纹饰,明显,网眼大小不一。
V.amoena(图1 D~F)极轴长30.0,赤道轴长37.6,P/E值为0.8。赤道面观为扁球形,极面观为三裂圆形。三孔萌发沟并延伸达两极,萌发沟细窄,少数花粉萌发沟中部有凸起。表面为穴状纹饰,较深。
V.sativa2505(图2 A~C)极轴长24.3,赤道轴长34.8,P/E值为0.7。赤道面观为扁圆形,极面观为三裂圆形。三孔萌发沟并延伸达两极,萌发沟较宽,沟中部有明显突起。表面为网状纹饰,较深,网眼大小不一。
V.sativa2560A(图2 D~F)极轴长20.5,赤道轴长25.8,P/E值为0.8。赤道面观为扁球形,极面观为三裂圆形。三孔萌发沟并延伸达两极,萌发沟细窄,沟中部有明显突起。表面为脑状纹饰,较深,纹饰层次分明。
V.sativa2560B(图3 A~C)极轴长21.8,赤道轴长25.6,P/E值为0.9。赤道面观为扁球形,极面观为三裂圆形。三孔萌发沟并延伸达两极,萌发沟深且宽。表面为片状纹饰,非常明显。
V.villosa(图3 D~F)极轴长10.3,赤道轴长9.0,P/E值为1.1。赤道面观为长球形,极面观为三裂圆形。三孔萌发沟并延伸达两极,萌发沟细窄,沟中部有明显凸起。表面为网状纹饰,较浅,网眼大小不一。
V.sativa2556(图4 A~C)极轴长30.5,赤道轴长47.4,P/E值为0.6。赤道面观为扁圆形,极面观为三裂圆形。三孔萌发沟并延伸达两极,萌发沟细窄,沟中部有细小凸起。表面为脑状纹饰,比较明显。
V.fabavar.minuta(图4 D~F)极轴长19.3,赤道轴长20.8,P/E值为0.9。赤道面观为扁球形,极面观为三裂圆形。三孔萌发沟并延伸达两极,萌发沟细窄,沟中部有明显凸起。表面为网状纹饰,比较模糊,网眼大小不一。
3 讨论
苏亚拉图等[12]认为,V.amoena花粉粒外壁为细网状纹饰或模糊,萌发沟中部有疣状凸起,而本研究观察发现,此物种的花粉粒外壁为穴状纹饰,萌发沟比较平滑(图1 D~F)。这8种植物花粉粒形态特征从以下方面进行识别和区分,即花粉的极轴长和赤道轴长,赤道面观,外壁纹饰。

图2 V.sativa 2505(A~C)和V.sativa 2560A(D~F)花粉粒形态Fig.2 Pollen morphology of V.sativa 2505(A~C)and V.sativa 2560A(D~F)

图3 V.sativa 2560B(A~C)和V.villosa(D~F)花粉粒形态Fig.3 Pollen morphology of V.sativa 2560B(A~C)and V.villosa(D~F)

图4 V.sativa 2556(A~C)和V.faba var.minuta(D~F)花粉粒形态Fig.4 Pollen morphology of V.sativa 2556(A~C)and V.faba var.minuta(D~F).
按照花粉粒极轴长将这8份种质从大到小排序,即V.sativa2556,V.amoena,V.fabavar.faba,V.sativa2505,V.sativa2560B,V.sativa2560A,V.fabavar.minuta,V.villosa(表2);同样,按照花粉粒赤道轴长可排为V.sativa2556,V.amoena,V.sativa2505,V.fabavar.faba,V.sativa2560A,V.sativa2560B,V.fabavar.minuta,V.villosa。表明,这8份种质花粉粒的极轴长与赤道轴长没有直接的相关性。根据花粉粒体积大小演化的一般趋势,即比较原始的植物花粉粒体积较大[20]。因此,可以根据这8个物种花粉粒的极轴长和赤道轴长推测它们的历史演化趋势。
通过对野豌豆属8种植物(1种已见报道)的花粉进行扫描电镜观察,既对已报道的结果进行验证,又同时获得了新材料的花粉形态学信息,对丰富野豌豆属植物孢粉学的信息做出了一定贡献。在已报道的V.amoena发现差异较大,本研究有明确的引种渠道和形态学分类结果,获得了清晰的花粉粒扫描电镜照片(图1 D~F);而前人仅在文中做了描述,并未提供图片。从而表明,对植物孢粉学观察结果进行验证是必要的。
有研究资料表明,可将花粉形态特征作为植物的分类依据[21]。V.amoena极轴长与V.sativa2505和V.villosa差异显著(P<0.05),推测二者亲缘关系较远;V.fabavar.faba与V.sativa2556和V.villosa的花粉在赤道轴长上差异显著,推测二者亲缘关系较远。这一结果有关学者通过分子标记已经证实[22-23]。花粉作为植物的保守器官,其形态特征具有可遗传性,同时具有科属的共同特征和种的特异性[24]。张庭龙等[25]认为花粉粒大小可能与植物生长环境有关,花粉形态要比花粉绝对大小更加稳定。Beck[26]指出,植物花粉在原始时期都是光滑且具有无定型的外壁,可以推测原始的野豌豆属植物花粉外壁都是光滑的,依据进化程度的不同,外壁纹饰也有明显差异,这为鉴定物种的特异性提供了依据[27]。长柔毛野豌豆外壁纹饰为网状纹饰,较浅,推测其进化上处于较原始的地位。V.sativa2560A和V.sativa2560B的外壁纹饰都非常明显,推测其处于较为进化的地位。此结果有关学者在野豌豆属植物的核型研究上已经得到证实[28]。本研究发现,依据植物花粉形态和外壁纹饰类型,可鉴定野豌豆属的共同特征和种的特异性。此外,还可根据花粉粒的极轴长、赤道轴长和萌发孔形态特征将这8份豌豆属植物材料区分开来。此外,根据所研究的8份野豌豆属植物材料花粉粒大小的演化趋势还可推测各个材料的进化程度甚至亲缘关系,为野豌豆属牧草系统分类学奠定基础,为其孢粉学相关研究积累信息。
[1] Liu Z P,Ma L C,Nan Z B,etal.Comparative transcriptional profiling provides insights into the evolution and development of the zygomorphic flower ofViciasativa(Papilionoideae)[J].PloS One,2013,8(2):e57338.
[2] 贺学礼,赵莉丽,谭志远,等.7种豆科植物花粉形态的研究[J].中国草地,1994(3):62-64.
[3] 姚雅琴,蒋选利,郁枫,等.豆科Leguminosae植物部分种花粉形态研究[J].西北农业大学学报,1990,18(2):78-82.
[4] Castiglione M R,Frediani M,Gelati M T,etal.Cytology ofViciaspecies X.karyotype evolution and phylogenetic implication inViciaspecies of the sectionsAtossa,Microcarinae,WiggersiaandVicia[J].Protoplasm,2011,248(4):707-716.
[5] Ball P W.ViciaL[A].In:Tutin T G,Heywood V H,Burges N A,etal.Flora Europaea[C].Cambridge:Cambridge University Press,1968:129-136.
[6] Castiglione M R,Frediani M,Ravalli C,etal.Cytological characterization ofViciaoroboidesWulfen in Jacq[J].Protoplasm,2009,236(1-4):21-27.
[7] Frediani M,Caputo P,Venora G,etal.Nuclear DNA contents,rDNAs and karyotype evolution in subgenusVicia:III.The heterogeneous sectionHypechusa[J].Protoplasm,2006,228(4):167-177.
[8] Frediani M,Caputo P,Venora G,etal.Nuclear DNA contents,rDNAs and karyotype evolution inViciasubgenusVicia:II.SectionPeregrinae[J].Protoplasm,2005,226(3-4):181-190.
[9] Maxted N.A phenetic investigation ofViciaL.subgenusVicia(Leguminosae,Vicieae)[J].Botanical Journal of the Linnean Society,1993,111(2):155-182.
[10] 夏振岱.中国野豌豆属的分类研究[J].植物分类学报,1996,34(4):421-433.
[11] 宋敏,于洪柱,娄玉洁,等.山野豌豆生物学特性及其利用[J].草业与畜牧,2011(4):5-6.
[12] 苏亚拉图,音扎布,哈斯巴根.内蒙古野豌豆属(ViciaL.)植物的花粉形态研究[J].内蒙古师大学报,1998,27(3):215-222.
[13] 苏亚拉图,哈斯巴根,音扎布.内蒙古野豌豆属-新变种及其花粉形态[J].内蒙古师大学报,2000,29(2):135-136.
[14] Erdtman G.Palynology[A].In:Preston R D.Advances in Botanical Research[C].London:Academic Press,1963:149-208.
[15] Grayum M H.Correlations between pollination biology and pollen morphology in the Araceae,wigh some implications for angiosperm evolution[A].In:Blackmore S,Ferguson I K.Pollen and Spores:Form and Function[C].London:Academic Press,1986:313-327.
[16] Nilsson S.The significance of pollen morphology in the Apocynaceae[A].In:Blackmore S,Ferguson I K.Pollen and Spores:Form and Function[C].London:Academic Press,1986:313-327.
[17] 卢玉飞,蒋建雄,艾辛,等.芒属部分类群花粉形态观察研究[J].草业学报,2012,21(6):151-158.
[18] 张彦妮,钱灿.12种百合属植物花粉形态扫描电镜观察[J].草业学报,2011,20(5):111-118.
[19] Punt W,Hoen P,Blackmore S,etal.Glossary of pollen and spore terminology[J].Review of Palaeobotany and Palynology,2007,143(1-2):1-81.
[20] 刘家熙,席以真,宁见长,等.中国紫草科破布木属花粉形态和外壁超微结构[J].植物学报,2001,43(9):893-898.
[21] 丁春帮,李强,李燕,等.重楼属9种5变种花粉活力与柱头可授性研究[J].草业学报,2009,18(4):61-66.
[22] 贺蓉,贺士元.华北地区野豌豆属的系统研究[J].西北植物学报,1994,14(4):303-306.
[23] Elena P,Tomooka N,Duncan A V,etal.Phylogeny ofVicia(Fabaceae)based on analysis of RAPDs and RFLP of PCR-amplified chloroplast genes[J].Genetic Resources and Crop Evolution,1999,46(2):149-161.
[24] 蓝盛银,徐珍秀.植物花粉剥离观察扫描电镜图解[M].北京:科学出版社,1996:42-45.
[25] 张庭龙,张启翔,谢松林.秦巴山及其毗邻地区8种野生百合孢粉学研究[J].西北农业学报,2010,19(1):144-146.
[26] Beck C B.被子植物的起源和早期演化[M].北京:科学出版社,1981:190-194.
[27] 王彦荣,南志标,聂斌,等.几种抗寒春箭筈豌豆新品系的形态特异性比较[J].草业学报,2005,14(2):28-32.
[28] 冉雪琴,朱晓彤,朱邦长.贵州野豌豆属植物的核型研究[J].草业科学,1997,14(1):5-9.
猜你喜欢
红蜻蜓·低年级(2024年6期)2024-07-08 09:22:17
儿童故事画报·自然探秘(2024年6期)2024-05-22 22:26:48
作物杂志(2021年6期)2021-12-17 07:20:24
植物研究(2021年5期)2021-03-02 03:53:10
草地学报(2017年1期)2017-09-14 09:10:20
西南农业学报(2016年5期)2016-05-17 05:42:43
东北林业大学学报(2015年2期)2015-03-10 03:11:32
军事文摘·科学少年(2015年2期)2015-01-26 14:43:32
—— 对极轴(下)
天文爱好者(2014年1期)2014-10-11 00:42:20
草食家畜(2010年4期)2010-03-20 13:22:26
