
青藏高原3种野豌豆光合生理特性的比较
2017-09-14 09:10刘玉萍周勇辉张晓宇
草地学报 2017年1期
刘玉萍,周勇辉,吕 婷,张晓宇,苏 旭
(青海师范大学 生命与地理科学学院 青藏高原药用动植物资源重点实验室 青海省自然地理与环境过程重点实验室青藏高原环境与资源教育部重点实验室,青海 西宁810008)
野豌豆属(Vicia)是豆科(Leguminosae)、野豌 豆族(Vicieae)一个比较重要的属,全世界约有200种,广泛分布于北半球温带至南半球温带以及东非等地,中国有43种5变种,主产于西北、华北和西南地区,尤以西北地区的青海省最多,有15种1变种,广泛生长于田边、路旁和荒地[1]。该属植物营养丰富,含有丰富的蛋白质,适口性好,在畜牧业生产上具有重要的经济价值;同时,在农业上可以作为牧草、绿肥、食用以及观赏用。其中,野豌豆属中的救荒野豌豆(Viciasativa)、山野豌豆(V.amoena)和三齿萼野豌豆(V.bungei)在青海省乃至整个青藏高原地区具有更加广泛的用途和经济生态价值,其不仅具有较高的饲用价值,被用作青饲、放牧、调制干草或青贮,是农牧业上的优良牧草,而且还可作为早春蜜源植物或水土保持植物,同时也是较为珍贵的药用植物[2]。
有关青藏高原高山植物光合生理特性的研究报道始于20世纪70年代[3]。目前,国内外学者已对青藏高原高山植物的光合生理特性作了大量的研究工作[4-10],然而,有关青藏高原豆科、野豌豆属植物光合生理特性的研究却至今未见报道。据此,本研究以青藏高原东北部自然生长的3种野豌豆属植物—救荒野豌豆、山野豌豆和三齿萼野豌豆为研究对象,系统探讨其光合色素含量、光合速率和叶绿素荧光参数等多种光合生理指标,并对这些光合生理指标的变化与参试物种的相关性、以及其对环境的适应性作进一步的分析,以期明确青藏高原地区野豌豆属植物光合生理指标的变化特征及其对逆境生态环境因子的适应机理,旨在为野豌豆属植物的人工栽培和开发利用提供生理学方面的理论依据。
1 材料与方法
1.1 材料
试验材料采自青藏高原东北部的青海省西宁市郊 区(E 101°46′,N 36°37′,海 拔 为 2280 m)。试验区为高原大陆性气候,年辐射总量为588~617kJ·cm-2,平均气压为746.2~778.7 hPa,年平均气温为2.8~3.5℃。2015年7月8-10日,将3种野豌豆属植物(每个物种采集8~10株)分别从试验点连根带土移栽至塑料花盆中(保证移栽后植株盆栽生长土壤环境与其生长地土壤环境相近),采回后在中国科学院西北高原生物研究所院内半遮阴环境中经短暂适应后(1~2d),于室内人工光源下进行光合速率及叶绿素荧光参数的测定。
1.2 方法
1.2.1 叶片厚度的测定 在移栽至花盆中的3种野豌豆属植物不同的植株上分别选取生长健康、长势一致、光照均匀的叶片,用电子数显外径千分尺(0~25,桂林广陆量具厂,中国)测定其叶片厚度,测定时避开主脉和主要次生叶脉,以植株叶片中下部2/3部位为测定部位,分左右测定,样本数n=70。
1.2.2 光合速率的测定 将移栽至花盆中的3种野豌豆属植株放置在无强光直接照射的半遮荫环境中适应1~2d,在植株上选取光照较一致并且长势较均一的健康叶片,在室内用Clark氧电极(LSZH,Hansatech公司,英国)分别测定0,45,90,180,360,450,900,1440,1800μmol·m-2·s-1光强下的净光合速率(Pn);测定时叶片在每个光强下平衡大约3~5min,每种植物的样本进行4~5次重复测定。表观量子效率AQY用0~150μmol·m-2·s-1范围内净光合速率的直线回归斜率来表示。
1.2.3 叶绿素和类胡萝卜素含量的测定 光合速率测定完成后,用电子天平(BS200S,Sartorius公司,德国)称量3种野豌豆属植物的叶片质量,并用叶面积仪(CI-203,CID公司,美国)测定叶面积,然后放入10mL体积比为45%乙醇、45%丙酮和10%蒸馏水的混合液中浸泡,于4℃遮光保存,约7 d后用紫外分光光度计(UV-1601,日本,导津公司)测定浸出溶液在663,645,440,505,652nm 波长下的吸光值。依照Arnon公式[11]计算单位叶面积叶绿素和类胡萝卜素的浓度和含量,并计算Chl a/b、叶黄素/叶绿素(A505/A652)值[9]。设置3次重复。
1.2.4 叶绿素荧光参数的测定 将3种野豌豆属盆栽植株放入植物培养实验室内暗适应约12h后,于次日清晨在避免自然光照射情况下,利用便携脉冲调制式荧光仪(FMS-2,Hansatech公司,英国)测定F0、Fm和光系统II(PSII)最大光化学效率(Fv/Fm);随后将从植株上摘取的叶片放入盛有少量蒸馏水的玻璃培养皿中,对叶柄进行适度剪切后保证叶柄基部浸入水中(为了避免叶片在人工光辐射下上表皮过分蒸腾引起气孔关闭,进而影响光合气体交换和电子传递过程,故在玻璃培养皿上用透光率为90%Luminar薄膜覆盖);将放有叶片的玻璃培养皿置于荧光灯培养架下,以光合有效辐射为450 μmol·m-2·s-1的光强进行照射,隔1h后取出叶片,用湿滤纸夹住叶柄基部,暗适应5min后测定Fv/Fm;约5h光照处理后测定作用光下的叶绿素稳态荧光(Fs)、最大荧光强度(Fm′)等参数;关闭培养架荧光灯,测定不同暗恢复时期的叶片荧光参数Fv/Fm。叶绿素荧光参数分别参照Oxborough和Baker[12]、Bilger和 Björkman[13]、Genty 等[14]方 法计算。每个物种的样本设置4个重复。
1.3 数据分析
数据用SPSS 17.0统计软件进行统计分析,3个野豌豆属物种间的差异采用单因素方差分析(one-way ANOVA),多重比较采用最小显著差异法(LSD),显著性水平设定为α=0.05,数据均以平均值和标准误(SE)表示;同时,利用Microsoft Excel制图。
2 结果与分析
2.1 叶片厚度
研究结果表明,救荒野豌豆的平均叶片厚度为0.1231mm,山野豌豆的为0.1956mm,三齿萼野豌豆的为0.2043mm。多重比较分析显示,3种野豌豆属植物的叶片厚度在救荒野豌豆与山野豌豆和三齿萼野豌豆间具有显著差异(P<0.05),而山野豌豆和三齿萼野豌豆间差异不显著(图1)。

图1 3种野豌豆属植物叶片厚度的比较Fig.1 Comparison of leaf thickness of three Vicia species
2.2 光合速率
在一定光强范围内,3种野豌豆属植物叶片的净光合速率随光量子通量密度(PFD)的增加而增加(图2)。在相同光量子通量密度下,救荒野豌豆叶片的净光合速率最高,为15.48μmol·m-2·s-1;其次是山野豌豆,为6.99μmol·m-2·s-1;三齿萼野豌豆的最低,为4.00μmol·m-2·s-1(表1)。同时,多重比较分析表明,救荒野豌豆和山野豌豆的净光合速率均显著大于三齿萼野豌豆的净光合速率(P<0.05),而救荒野豌豆和山野豌豆间的净光合速率无显著差异(表1),这说明前两者比三齿萼野豌豆具有更高的光合能力。然而,当光量子通量密度达1400μmol·m-2·s-1时,3种野豌豆属植物叶片的Pn均已达到饱和,当光量子通量密度高于1400μmol·m-2·s-1时,救荒野豌豆叶片的Pn开始出现下降,产生光抑制现象,从而引起Pn的降低;而山野豌豆和三齿萼野豌豆植物叶片的Pn此时并未出现下降,表明没有光抑制现象发生(图2)。

图2 3种野豌豆属植物叶片净光合速率对光量子通量密度的响应Fig.2 Response of net photosynthesis rate(Pn)to photon flux density(PFD)of three Vicia species
3种野豌豆属植物叶片的表观量子效率AQY与Pn的变化趋势和差异性基本一致(表1),即在相同光量子通量密度下,救荒野豌豆叶片的AQY最高,为0.06μmol·μmol-1;山野豌豆叶片的AQY次之,其值为0.05μmol·μmol-1;三齿萼野豌豆叶片的AQY最低,为0.03μmol·μmol-1;并且,救荒野豌豆和山野豌豆的AQY与三齿萼野豌豆存在显著差异(P<0.05),而救荒野豌豆和山野豌豆之间差异不显著(表1);然而,3种野豌豆属植物叶片光补偿点Ic的变化趋势与AQY的变化趋势相反,表现为救荒野豌豆(23.43μmol·m-2·s-1)<山野豌豆(55.48μmol·m-2·s-1)<三齿萼野豌豆(63.42μmol·m-2·s-1),并且光补偿点在3种野豌豆属植物相互间均存在显著差异(P<0.05),这表明3种野豌豆叶片的光补偿点与其净光光合速率呈负相关关系(表1)。此外,在3种野豌豆属植物中,山野豌豆的暗呼吸速率Rd最高,为-2.52μmol·m-2·s-1;其次是三齿萼野豌豆,为-2.02μmol·m-2·s-1;救荒野豌豆最低,仅为1.55μmol·m-2·s-1。同时,多重比较分析显示,3种野豌豆属植物中仅有救荒野豌豆与三齿萼野豌豆之间的暗呼吸速率存在显著差异(P<0.05),而在救荒野豌豆与山野豌豆以及山野豌豆与三齿萼野豌豆之间的暗呼吸速率差异均不显著(表1)。

表1 3种野豌豆属植物光合作用光响应参数比较Table 1 Comparison of photosynthetic parameters of three Vicia species
2.3 叶绿素和类胡萝卜素含量
由表2可知,3种野豌豆属植物叶片中总叶绿素(Chlt)和类胡萝卜素(Car)含量按物种救荒野豌豆、山野豌豆和三齿萼野豌豆的顺序依次升高,而叶绿素a/b比率(Chla/b)则相反,即三齿萼野豌豆的最低,山野豌豆的较高,救荒野豌豆的最高;同时,3种野豌豆之间叶黄素/叶绿素(A505/A652)值差异不显著。多重比较分析结果显示,三齿萼野豌豆与救荒野豌豆以及三齿萼野豌豆与山野豌豆间的总叶绿素含量存在显著差异,并且三齿萼野豌豆和救荒野豌豆间的叶绿素a/b也具有显著差异(P<0.05);相反,救荒野豌豆与山野豌豆间的总叶绿素含量、山野豌豆与救荒野豌豆以及山野豌豆与三齿萼野豌豆间的叶绿素a/b、以及3种野豌豆属植物相互间的叶黄素/叶绿素及类胡萝卜素均无显著差异。
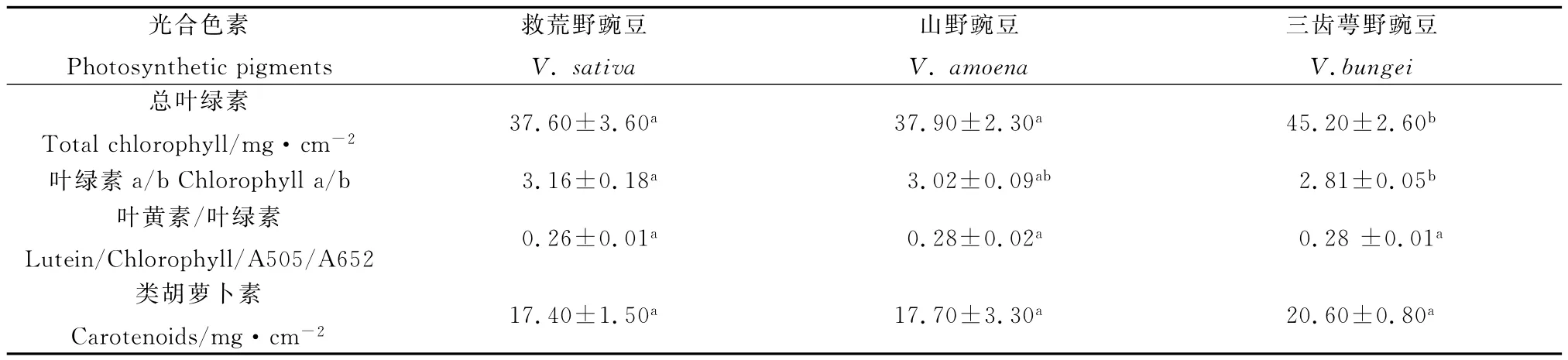
表2 3种野豌豆属植物叶绿素和类胡萝卜素含量的比较Table 2 Comparison of chlorophyll and carotenoid contents of three Vicia species
2.4 叶绿素荧光参数的变化
由图3和图4可知,3种野豌豆属植物叶片的光系统II实际光化学量子效率(ФPSII)和光化学猝灭系数(qp)随光量子通量密度的增加呈明显的下降趋势。总体而言,山野豌豆的光系统II实际光化学量子效率和光化学猝灭系数显著高于救荒野豌豆和三齿萼野豌豆,其中三齿萼野豌豆最低。
由图5可知,随光量子通量密度的增加,3种野豌豆属植物叶片的非光化学猝灭系数(NPQ)呈现上升趋势,并且救荒野豌豆叶片的非光化学猝灭系数最高,山野豌豆的次之,三齿萼野豌豆的最低。

图3 3种野豌豆属植物光系统II实际光化学速率(ФPSII)对光量子通量密度的响应Fig.3Response of actual photochemical efficiency of PSII(ФPSII)to photon flux density(PFD)of three Vicia species
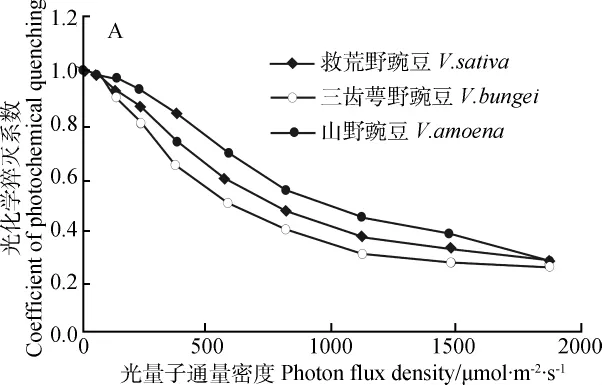
图4 3种野豌豆属植物光化学猝灭系数(qp)对光量子通量密度的响应Fig.4 Response of coefficient of photochemical quenching(qp)to photon flux density(PFD)of three Vicia species

图5 3种野豌豆属植物非光化学猝灭系数(NPQ)对光量子通量密度的响应Fig.5 Response of coefficient of non-photochemical quenching(NPQ)to photon flux density(PFD)of three Vicia species
由图6可知,在一定光强范围内,3种野豌豆属植物叶片的电子传递速率(ETR)随光量子通量密度的增加而增加,当光量子通量密度达到约600 μmol·m-2·s-1时,救荒野豌豆、山野豌豆和三齿萼野豌豆叶片的电子传递效率都到最大值;随后,随光量子通量密度的继续增大,除救荒野豌豆叶片的电子传递效率基本不变外,山野豌豆和三齿萼野豌豆叶片的电子传递效率呈现下降趋势;当光量子通量密度接近2000μmol·m-2·s-1时,救荒野豌豆和山野豌豆叶片的电子传递效率大致相等。总体看来,叶片电子传递效率的大小顺序为山野豌豆>救荒野豌豆>三齿萼野豌豆。

图6 3种野豌豆属植物电子传递速率(ETR)对光量子通量密度的响应Fig.6 Response of electron transfer rate(ETR)to photon flux density(PFD)of three Vicia species
3 讨论与结论
光合作用是植物对逆境反应最为敏感的生理过程之一[3]。西宁市位于青藏高原东北部,气温低、积温少、气压低、太阳辐射强,因而生长于该地区的植物,由于长期受环境中多种逆境生态因子的胁迫,在形态和生理生化上必定产生一定的进化适应方式[15-17],尤其在光合生理特性上发育了一套具有适应该环境的特殊生理机制。高山植物的光合生理特性反映了其对高原特殊自然环境条件的响应和适应;响应反映在植物生理功能对短期环境变化的反应,适应主要指在长期的特定环境中植物所形成的形态、结构以及生理功能的变化[6]。
3.1 叶片厚度变化趋势分析
苏旭和刘玉萍[9-10]研究发现,高山植物叶片厚度的增加可有效防止强光辐射对植物光合系统的损伤。3种野豌豆属植物中,三齿萼野豌豆和山野豌豆的叶片明显厚于救荒野豌豆,说明前两者更能有效防止青藏高原地区强光辐射对其叶片光合系统的破坏,在一定程度上提高了其对强光的适应能力,从而导致他们在青藏高原地区具有相对较广的地理分布范围。
3.2 光合作用的光响应特性分析
青藏高原地区植物由于长期受低温、低气压等多种逆境生态因子的影响,加之强辐射下植物常常存在一定程度的光合抑制现象,因而他们的光合作用表观量子效率往往偏低[6]。表观量子效率AQY是光合作用中光能转化最大效率的一种度量,可以反映光合机构中光合机能的变化[18]。我们的研究表明,在一定光强范围内随着光量子通量密度(PFD)的增加,3种野豌豆属植物叶片的表观量子效率AQY都随之增加;三齿萼野豌豆的表观量子效率显著低于救荒野豌豆和山野豌豆,说明三齿萼野豌豆没有其他2种野豌豆对CO2的羧化能力高,并且这种现象与师生波等[6]对青藏高原几种典型高山植物光合特性的研究结果类似。高山植物AQY较低,表明植物吸收的日光用于羧化CO2的效率低,可能与高原地区的强辐射和低CO2分压有关,同时存在一定的光合作用光抑制现象[19]。据此,我们推测三齿萼野豌豆的AQY较低也许并非是由于强辐射、低CO2分压及光合作用光抑制造成的,而可能是由于其叶片中活化的Rubisco酶含量较少所导致的,这种含量较少的酶将会使三齿萼野豌豆对CO2的羧化限制程度增大,进而致使其AQY小于救荒野豌豆和山野豌豆。同样,在一定光强范围内,随着光量子通量密度的增加,3种野豌豆属植物叶片的净光合速率也随之增加;并且,在相同光量子通量密度下,救荒野豌豆叶片的净光合速率高于山野豌豆和三齿萼野豌豆,表明救荒野豌豆比山野豌豆和三齿萼野豌豆具有更高的光合能力,这种结果与师生波等[6]对高山植物麻花艽(Gentianastraminea)的研究结果一致。我们认为,在同一高山环境条件下,三齿萼野豌豆比其他2种野豌豆属植物具有更低的净光合速率可能是由于低温与光照相“偶联”致使其叶片气孔导度比其他2个物种的气孔导度降低的更快所导致的,可能是其在高山地区分布较广、适应性较强的一个主要原因,也是其对低温与强太阳辐射等逆境生态因子的一种生理性适应和保护措施。
3种野豌豆属植物光合生理指标的变化,除体现在净光合速率(Pn)和表观量子效率(AQY)上,还体现在光补偿点(Ic)和暗呼吸速率(Rd)方面。高山植物光补偿点和暗呼吸速率与海拔高低关系的研究表明,随着海拔的升高,高山植物的光补偿点和暗呼吸速率均降低,认为光补偿点的降低是高山植物对强辐射生态环境的一种适应,暗呼吸速率的降低可能与温度降低有关,是高山植物抵抗寒冷的一种生理性适应[9,19]。本研究表明,3种野豌豆属植物在同一外部高山生态环境条件下,救荒野豌豆的光补偿点最低、三齿萼野豌豆的光补偿点最高,山野豌豆的暗呼吸速率最高、救荒野豌豆的暗呼吸速率最低,说明救荒野豌豆相比山野豌豆和三齿萼野豌豆,具有更好的抵抗强辐射和低温的能力,更易在极端环境生态条件下生存和生长。
3.3 光合色素含量变化分析
许大全[20]认为,光合色素在植物光合作用过程中具有光能吸收、传递和转化的作用。在本研究中,3种野豌豆属植物叶片中总叶绿素含量按照救荒野豌豆、山野豌豆和三齿萼野豌豆的顺序依次升高,并且3种野豌豆之间总叶绿素含量无显著差异(表2);3种野豌豆属植物叶片中,三齿萼野豌豆的叶绿素a/b比值最低,表明该物种叶绿素a的含量较低,叶绿素b含量较高,说明三齿萼野豌豆叶片中捕光色素系统发达[21],相比其他2个物种,三齿萼野豌豆对光能的捕获能力更强,亦表明在一定范围内其更能适应弱光环境[22]。类胡萝卜素是一种重要的保护性物质,具有辅助捕光色素捕获光能的功能,且对植物体内产生的活性氧自由基具有清除作用,从而可有效减轻活性氧对植物体的毒害,这样植物体进而能够维持较高水平的抗氧化能力和光合作用的高效进行[23]。3种野豌豆属植物中,三齿萼野豌豆叶片中类胡萝卜素的含量最高,救荒野豌豆中的含量最低。孙小玲等[23]认为,海拔越高,太阳辐射越强,紫外线辐射也越强,三齿萼野豌豆叶片中类胡萝卜素含量相对较高是其更能适应当地环境因素的重要体现,这也许是其在青藏高原地区分布相对较广的一个重要原因。此外,我们的研究还发现,3种野豌豆属植物叶片中叶绿素含量与其净光合速率呈负相关关系,这与先前某些学者[19-20]的观点一致。
3.4 叶绿素荧光参数变化分析
光系统II实际光化学速率反映PSII反应中心部分关闭时植物实际原初光能捕获效率,表示光化学反应消耗的能量比例,其值大小可以反映PSII反应中心的开放程度。研究发现,3种野豌豆属植物叶片的光系统II实际光化学量子效率(ФPSII)随着光量子通量密度的增加而下降,在稳定作用光强下山野豌豆的光系统II实际光化学量子效率明显高于救荒野豌豆和三齿萼野豌豆,尤其是三齿萼野豌豆(图3),表明3种野豌豆对于光能的利用存在一定差异,山野豌豆吸收的光能用于光化学过程的能力大于救荒野豌豆和三齿萼野豌豆,其中三齿萼野豌豆的最差,说明其更能适应强光环境,光抑制现象没有其他两个物种明显。
王强等[24]认为,过剩激发能的耗散对于光能的吸收和电子传递具有关键作用,并且对于防止光合器官的光抑制也至关重要。荧光猝灭是过剩光能耗散的一种重要形式,包含光化学荧光猝灭和非光化学荧光猝灭两种类型。光化学猝灭是光系统II天线色素吸收的光能中用于光化学电子传递的份额,与光系统II电子传递和初始电子受体QA的氧化还原有关[25],一定程度上反映光系统II反应中心的开放程度;非光化学猝灭(NPQ)是评价植物耗散过剩激发能的重要荧光参数,反映的是PSII天线色素吸收的光能不能用于光合电子传递而以热的形式耗散掉的激发能部分[26]。当PSII反应中心天线色素吸收的光能超过所能利用的量时,若不及时地耗散掉这部分过剩激发能,将对光合机构造成失活甚至破坏,故非光化学猝灭是植物的一种自我保护机制,对光合机构具有一定的保护作用[27]。在一定光强范围内,随着光量子通量密度的增加,3种野豌豆属植物的光化学猝灭系数呈明显的下降趋势;在恒定光强条件下,光化学猝灭系数在山野豌豆、救荒野豌豆和三齿萼野豌豆中依次呈降低趋势。光化学猝灭系数愈大,表明PSII反应中心的开放程度愈高[28],即山野豌豆PSII反应中心的开放程度在3种植物中是最大的;并且,3种野豌豆属植物的电子传递速率(ETR)亦可证明山野豌豆的电子传递速率明显高于其他另外2种野豌豆的电子传递速率。相反,研究还发现3种野豌豆属植物的非光化学猝灭系数(NPQ)随光量子通量密度的增加而增加,并且救荒野豌豆的非光化学猝灭系数最高、山野豌豆的次之、三齿萼野豌豆的最低,说明三齿萼野豌豆比救荒野豌豆和山野豌豆具有更强的将所捕获的光能有效的用于光合作用的能力,其热耗散能力较低,因此这也进一步表明三齿萼野豌豆较之山野豌豆和救荒野豌豆可能更适应于在青藏高原东北部的稍低海拔农业区大范围的引种栽培。
另外,表观光合电子传递效率(ETR)可以反映1,5—二磷酸核酮糖羧化氧化酶(Rubisco)的活性,包括氧化作用和羧化作用[14]。3种野豌豆属植物的表观光合电子传递速率(ETR)在一定光强范围内随光量子通量密度的增加而增加,当光量子通量密度达到约600μmol·m-2·s-1时,救荒野豌豆、山野豌豆和三齿萼野豌豆叶片的电子传递效率达到最高;从总体变化趋势看,山野豌豆的表观光合电子传递速率最大,三齿萼野豌豆的表观光合电子传递速率最小,救荒野豌豆的表观光合电子传递速居于前两者之间,这种变化趋势与光化学猝灭系数在3种野豌豆属植物中的变化趋势相吻合。
猜你喜欢
发明与创新(2021年9期)2021-12-06
——以“野豌豆”为例的课程资源建设初探
好家长(2021年4期)2021-12-01
科教新报(2021年9期)2021-05-17
科学(2020年1期)2020-08-24
河北画报(2020年19期)2020-06-16
——大别山野豌豆概述※
特种经济动植物(2019年9期)2019-10-29
种子(2017年2期)2017-12-02
北京信息科技大学学报(自然科学版)(2016年6期)2016-02-27
影像科学与光化学(2014年5期)2014-03-11
影像科学与光化学(2014年5期)2014-03-11
