
Loss of Function of Ferredoxin2 Can PromoteFlowering in Arabidopsis thaliana
2013-04-23 09:05,,,,,,
中山大学学报(自然科学版)(中英文) 2013年3期
, , , , , ,
(State Key Laboratory of Biocontrol// Guangdong Key Laboratory of Plant Resources, School of Life Sciences, Sun Yat-sen University, Guangzhou 510275, China )
Flowering is the turning point from the vegetative stage to the reproductive stage in the growth process of plants. InArabidopsisthaliana, the flowering time is regulated by many different pathways: gibberellin, autonomous, vernalization, and light-dependent pathways[1-2].
Plants utilize light as the signal to determine the timing of flowering. InArabidopsisthaliana, there are a lot of various photoreceptors to monitor light quality and quantity. These photoreceptors can perceive light to regulate the flowering time[3]. InArabidopsis, these photoreceptors include the red/far-red light-absorbing phytochromes (PHYs)( PHYA to PHYE)[3-5]and the blue/UV-A light-absorbing cryptochromes(CRYs) (CRY1 and CRY2)[6-9], phytochromes and cryptochromes work together to involve in regulating flowering[5,10]. In the previous studies, we have known the far-red light can promote flowering[5, 11, 12]and the far-red light photoreceptor mutant——phyAmutant displays a late-flowering phenotype when grown both under long-day and short-day conditions[10]. In contrast, red light can delay flowering[5, 11-13]and the red light photoreceptor mutant——phyBmutant displays an early-flowering phenotype when grown both under long-day and short-day conditions[10]. Moreover, thephyAphyBdouble mutant also displays an early-flowering phenotype when grown under long-day and short-day conditions[10]. So it could be concluded that the impairment in the physiology function of phytochromes can promote flowering in the overall effect.
In plants, to sense and harvest the light signals, the photoreceptors must maintain some structures under the participation of the chromophores. In plants, open chain tetrapyrroles can function as the chromophores of light-sensing phytochromes[14]. To synthesize tetrapyrrole chromophores, the heme is first linearized by heme oxygenase to form the open-chain tetrapyrrole intermediate ——biliverdin IXα(BV). BV is subsequently reduced by phytochromobilin (PФB) synthase ——HY2, which is a ferredoxin-dependent bilin reductase to produce the PФB chromophore for light-sensing phytochromes[15-16]. Therefore HY2 plays very important role in the formation of phytochromes and the physiology function of phytochromes. Moreover, thehy2 mutant also displays the early-flowering phenotype[17], this result further supports the conclusion that the impairment of the physiology function of phytochromes can promote flowering in the overall effect.
Ferredoxins(Fds) are the major donor systems for electrons to many various receptor systems in plastids. In photosynthetic organisms, Fds play roles not only in electron transfer system of photosynthesis but also in many redox reactions mediated by several oxidoreductases, such as sulfite reductase,nitrite reductase, glutamate synthase, and ferredoxin:thioredoxin reductase[18]. InArabidopsisthaliana, the PФB synthase ——HY2 has been shown to require ferredoxins (Fds) as the electron donors for reduction reaction[15-16].
Here, we found that a Ds-T-DNA insertion line ofArabidopsisthalianafor the gene encoding the most major ferredoxin (Fd2, At1g60950) can promote flowering in the process of growth both under long-day and short-day conditions. We show in this report that the loss of AtFd2functioncan promote flowering, AtFd2 can interact with AtHY2 in the chloroplast, and the loss of AtFd2 function can impair the responses mediated by phytochromes. These results implicate that the loss of AtFd2 can promote flowering by impairing the physiology function of phytochromes.
1 Materials and methods
1.1 Plant materials
For all experiments shown in this work,Arabidopsisthalianaplants of the ecotype Noessen WT andFd2-KOwere sent by Renate Scheibe and determined as described[17].
1.2 Flowering phenotype studies
The Noessen WT andFd2-KOArabidopsisthalianaseeds were germinated on soil and grown under long-day conditions (16h light/8h dark) or short-day conditions (8 h light/16 h dark) illuminated by white light at a fluence rate of 120μmol/m2/s. The number of days from germination to bolting, the number of rosette leaves when bolting, and the number of cauline leaves at maturity were scored.
1.3 BIFC study
Plasmid HY2-YN was made by cloning theAtHY2 fragment without the stop codon fromArabidopsiscDNA with the primers 5′-AGTCGACATGGCTTTATCAATGGAGT-3′ and 5′-TTACTCGAGGCCGATAAATTGTCCT-3′ and inserting into the pUC-SPYNE(YN) multiple cloning sites (MCS) withSalⅠandXhoⅠ.
Plasmid Fd2-YC was made by cloning theAtFd2 fragment without the stop codon fromArabidopsiscDNA with the primers 5′-TAGGATCCATGGCTTCCACTGCTCT-3′ and 5′-GCCCTCGAGAACAATGTCTTCTTCTT-3′ and inserting into the pUC-SPYCE(YC) multiple cloning sites (MCS) withBamHⅠandXhoⅠ.
Plasmids bZIP63-YN and bZIP63-YC were made by cloning thebZIP63 fragment without the stop codon fromArabidopsiscDNA and inserting into the pUC-SPYNE and pUC-SPYCE multiple cloning sites (MCS) withBamHⅠandXhoⅠ.
Construct pairs of bZIP63-YN and bZIP63-YC, HY2 -YN and Fd2-YC, pUC-SPYCE(YC) and HY2-YN, pUC-SPYNE (YN) and Fd2-YC, YN and YC were transiently expressed in rice protoplasts isolated from 9-day-old rice seedlings. Fluorescence was observed using an Olympus fluorescent microscope and visualized with Olympus DP2-BSW software.
1.4 Hypocotyl Measurements
Arabidopsisthalianaseeds were surface-sterilized in 50% bleach for 15 min and washed five times with sterile ddH2O before plating on a 1?2 MS plate with 0.8% (w) Agar. The seeds were vernalized in the dark at 4 ℃ for 4 days. Germination was induced with a 4 hr white light treatment. The seeds were followed grown at 23 ℃ in a LED chamber (Percival Scientific, Perry, IA) under dark and certain light conditions (red or far-red light ) for 6 days. For both photograph and hypocotyl length measurements, 6-day-old seedlings were photographed using a camera, and hypocotyl length was measured using ruler.
2 Results
2.1 Loss of AtFd2 can promote flowering in Arabidopsis thaliana
We found that theFd2-KOmutant flowered earlier than wild type both under long-day (16h light/8h dark) and short-day (8 h light/16h dark) conditions (Figure 1A and 2A). In order to further support the phenotype, we chose three most common physiological indexes(the number of days from sowing to bolting, the number of rosette leaves when bolting, and the number of cauline leaves at maturity) to score. These physiological indexes could reflect the period of flowering, generally speaking, the numerial number was smaller, the flowering was earlier. The results showed that the number of days from sowing to bolting, the number of rosette leaves, and cauline leaves ofFd2-KOmutant were all smaller than that of wild type (Figure 1B-D and Figure 2B-D). So we confirmed that the loss of AtFd2 can promote flowering inArabidopsisthaliana, and the loss of AtFd2 mayinvolve in the regulation of flowering inArabidopsisthalianathrough some pathway.
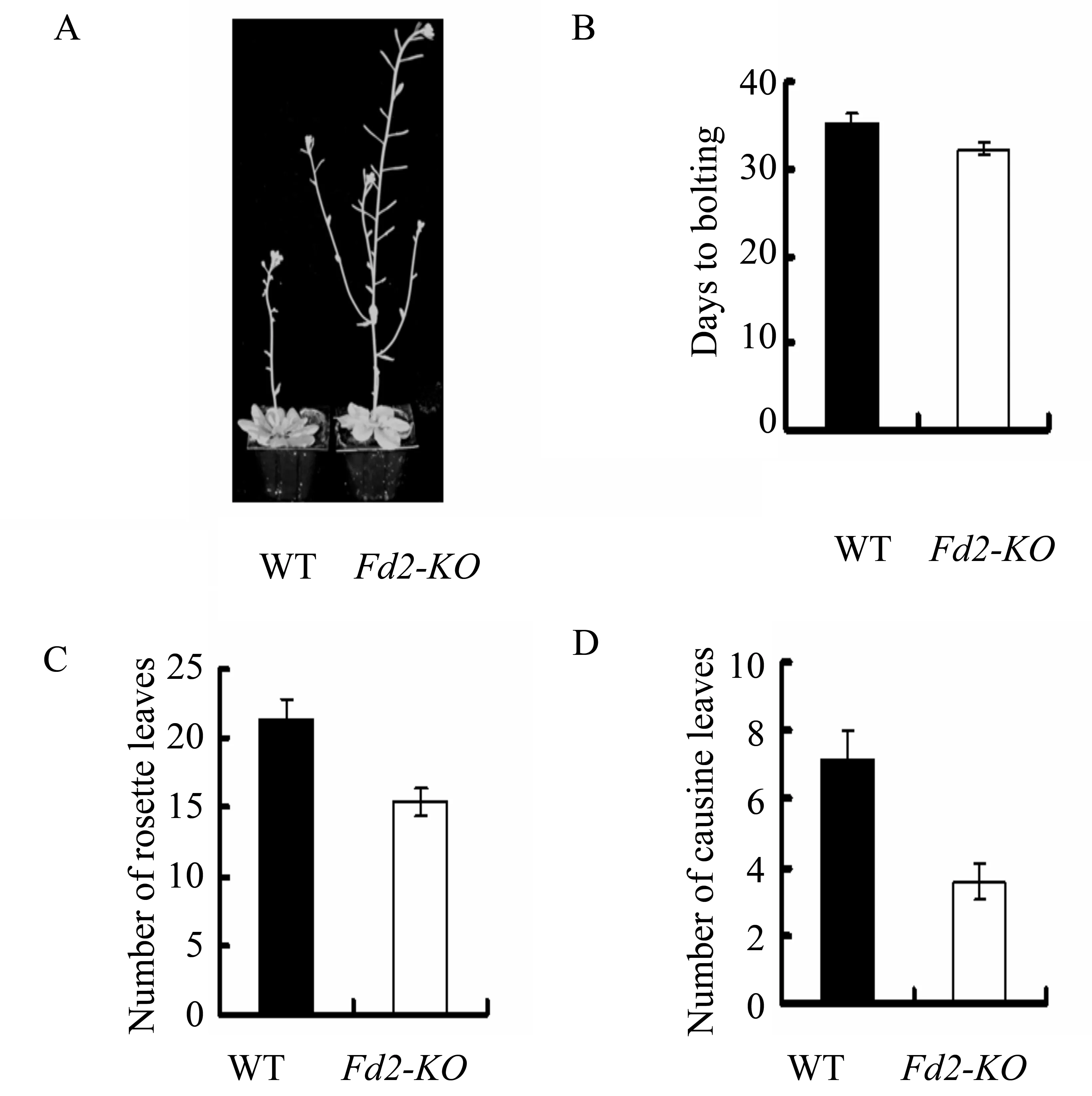
Fig.1 Phenotypes of the Noessen WT and Fd2-KO Arabidopsis plants under long-day conditionsA:Phenotypes of the Noessen WT and Fd2-KO Arabidopsis plants under long-day conditions;B:The number of days from sowing to bolting of the Noessen WT and Fd2-KO Arabidopsis plants under long-day conditions;C: The number of rosette leaves when bolting of the Noessen WT and Fd2-KO Arabidopsis plants under long-day conditions;D:The number of cauline leaves when bolting of the Noessen WT and Fd2-KO Arabidopsis plants under long-day conditions(Thirty plants of Noessen WT or Fd2-KO Arabidopsis thaliana were calculated, three independent experiments)

Fig.2 Phenotypes of the Noessen WT and Fd2-KO Arabidopsis plants under short-day conditionsA:Phenotypes of the Noessen WT and Fd2-KO Arabidopsis plants under short-day conditions;B:The number of days from sowing to bolting of the Noessen WT and Fd2-KO Arabidopsis plants under short-day conditions;C:The number of rosette leaves when bolting of the Noessen WT and Fd2-KO Arabidopsis plants under short-day conditions;D:The number of cauline leaves when bolting of the Noessen WT and Fd2-KO Arabidopsis plants under short-day conditions(Thirty plants of Noessen WT or Fd2-KO Arabidopsis thaliana were calculated, three independent experiments)
2.2 AtFd2 interacts with AtHY2 in the chloroplast
In order to further confirm whether AtFd2 interacts with AtHY2, we created the plasmid for expression ofAtHY2 fused to the N-terminal yellow fluorescence protein (YN) under control of the 35S-CaMV promoter and the plasmid for expression ofAtFd2 fused to the C-terminal yellow fluorescence protein (YC) under control of the 35S-CaMV promoter. And we transiently expressed HY2-YN and Fd2-YC together in the rice protoplasts. The fluorescence emission of YFP was monitored in rice protoplasts using an Olympus fluorescence microscope. We found that co-expression of the HY2-YN and Fd2-YC fusion proteins in rice green tissue protoplasts produced obvious YFP signals in the chloroplast (Figure 3D). This result is in agreement with several previous studies that AtHY2 and AtFd2 are localized to the chloroplast. And the result confirms that AtHY2 physically interacts with AtFd2. As the negative controls, co-expression of HY2-YN with empty pUC-SPYCE(YC) vectors, empty pUC-SPYNE(YN) with Fd2-YC vectors, or two empty vectors pUC-SPYNE(YN) with pUC-SPYCE(YC) produced no BiFC fluorescence (Figure 3A-C). As a positive control, the bZIP63-YN and bZIP63-YC fusion proteins in rice green tissue protoplasts produced obvious YFP signals in the nucleus (Figure 3E).
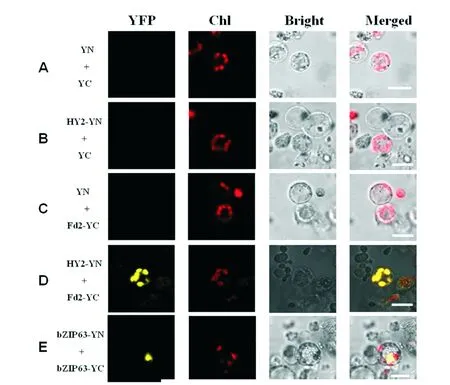
Fig.3 AtFd2 interacts with AtHY2 in the chloroplastConstruct pairs of bZIP63-YN and bZIP63-YC, HY2 -YN and Fd2-YC, pUC-SPYCE(YC) and HY2-YN, pUC-SPYNE (YN) and Fd2-YC, YN and YC were transiently co-expressed in rice green tissue protoplasts. BiFC fluorescence was indicated by the YFP signal. Individual and merged images of YFP and chlorophyll autofluorescence (Chl) as well as bright field images of protoplasts were shown. Scale bars = 10 μm
2.3 Fd2-KO mutants are impaired in the responses mediated by phytochromes
In order to study whetherFd2-KOMutants promote flowering by impairing the responses to both red light and far-red light mediated by phytochromes, we first examined the response to red light mediated by phytochromes inFd2-KOseedlings by measurement of hypocotyl lengths of 6-day-old seedlings compared to wild type grown under continuous red light. In dark conditons,Fd2-KOseedlings had shorter hypocotyl lengths than wild type since the loss of AtFd2 inhibited the normal growth of seedling. But the gap of the hypocotyl length betweenFd2-KOseedlings and wild type seedlings reduced with the increasement of the red light intensity(Figure 4A). Moreover,Fd2-KOseedlings had longer hypocotyl length than wild type seedlings under high red light intensity(Figure 4B).
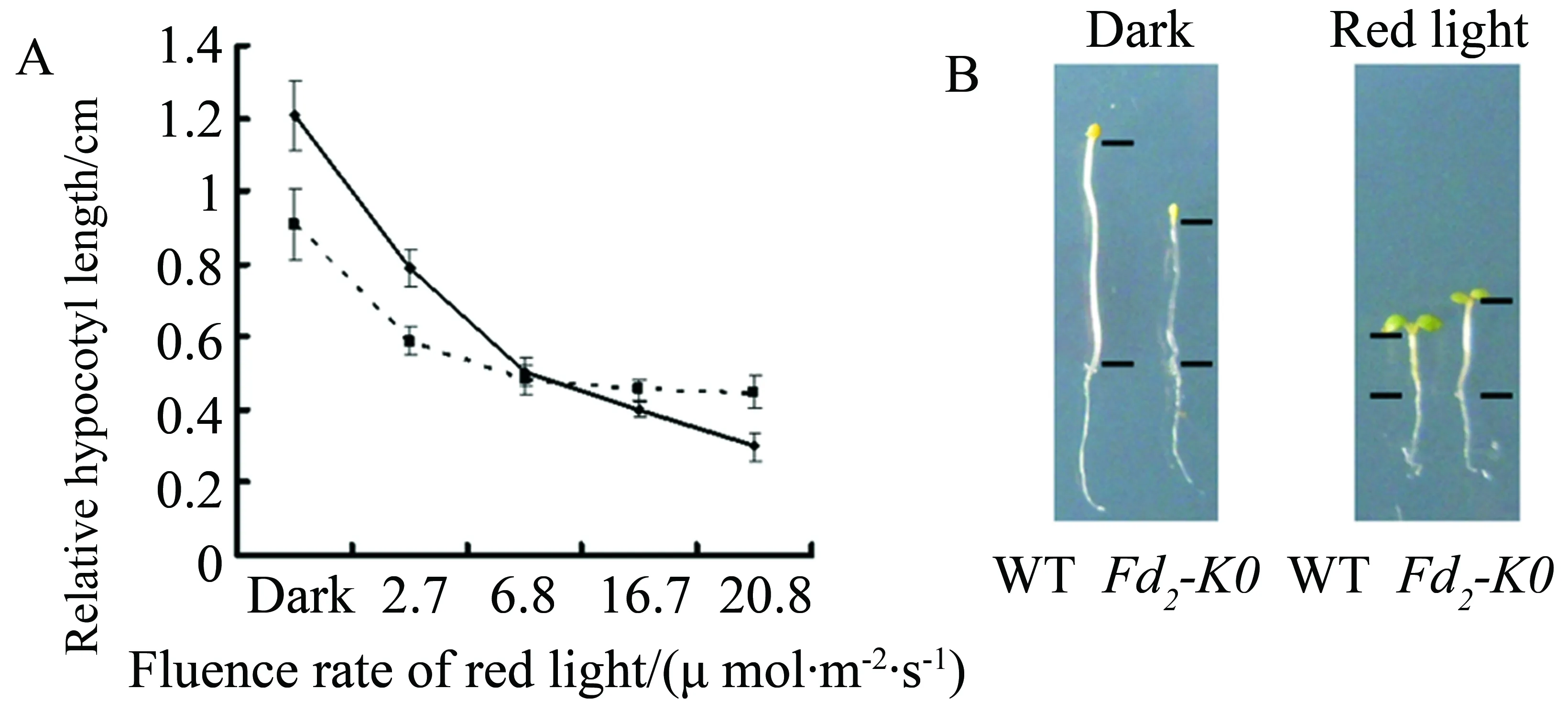
Fig.4 Phenotypes of the Noessen WT and Fd2-KO Arabidopsis seedlings under red light conditions.A:Relative hypocotyl length of 6-day-old grown Noessen WT(solid line), Fd2-KO (dotted line) Arabidopsis seedlings under different fluence of red light and dark conditions; B:Phenotypes of the 6-day-old grown Noessen WT and Fd2-KO Arabidopsis seedlings under red conditions(20.8 μmol·m-2·s-1)and dark conditions(Thirty plants of Noessen WT or Fd2-KO Arabidopsis thaliana were calculated, three independent experiments)
The response to far-red light mediated by phytochromes was also examined inFd2-KOseedlings by measurement of hypocotyl lengths of 6-day-old seedlings grown under continuous far-red light compared to wild type. The result was similar with the result of red light(Figure 5A and 5B). These results indicate thatFd2-KOMutants are impaired in the responses mediated by phytochromes and implicate that the loss of AtFd2 may promote flowering by impairing the physiology function of phytochromes.
3 Discussion
Based on the phenotype ofFd2-KOmutants, which flowered earlier than wild type both under long-day and short-day conditons, we proposed the possible pathway AtFd2 regulates in flowering .
The previous studies have shown that phytochromes are involved in the regulation of flowering. The far-red light photoreceptor-PHYA and the red light photoreceptor-PHYB are the most important phytochromes inArabidopsisthaliana. Based on the effect of far-red /red light and the phenotype ofphyA、phyB、phyAphyBmutants[5, 10-13], we may come to the conclusion that the impairment of the physiology function of phytochromes can promote flowering in the overall effect.
And we learned that one of the most important enzymes in the synthesis of chromophore for light-sensing phytochromes ——HY2, which is ferredoxin-dependent and requires ferredoxins (Fds) as the electron donors to perform its normal physiological function[15-16]. Moreover, thehy2 mutant also flowered early[18], this result further supported the conclusion that the impairment of the physiology function of phytochromes can promote flowering in the overall effect.
AtFd2 is the most important ferredoxin inArabidopsisthaliana, so we guessed that the loss of AtFd2 may promote flowering by impairing the physiology function of phytochromes.
We demonstrated that AtFd2 can interact with AtHY2 in chloroplast.Though the previous studies have proven that AtHY2 requires AtFd2 as the electron donors through the biochemical and structural studies, our results first prove that AtFd2 can interact with AtHY2 in vivo through BIFC.The results further support that AtHY2 is ferredoxin-dependent and may require AtFd2 as the electron donors for double bond reductions.
In conclusion, all the results implicated that the loss of AtFd2 may promote flowering by impairing the physiology function of phytochromes. However, because of the difference between the physiology progress mediated by phytochromes and the flowering progress, we can identify the genetic regulation relationship between AtFd2 and phytochromes to further confirm our conclusions in the following research.
[1] JAEGER K E, GRAF F. The control of flowering in time and space[J]. J Exp Bot, 2006, 57(13): 3415-3418.
[2] KOMEDA Y. Genetic regulation of time to flower inArabidopsisthaliana[J]. Annu Rev Plant Biol,2004, 55: 521-535.
[3] QUAIL P H. Phytochrome photosensory signalling networks[J]. Nat Rev Mol Cell Biol,2002, 3(2): 85-93.
[4] CHEN M, GALVAO R M, et al. Arabidopsis HEMERA/pTAC12 initiates photomorphogenesis by phytochromes[J]. Cell, 2010,141(7): 1230-1240.
[5] ENDO M, NAGATANI A. Flowering regulation by tissue specific functions of photoreceptors[J]. Plant Signal Behav, 2008, 3(1): 47-48.
[6] BRIGGS W R, CHRISTIE J M.Phototropins 1 and 2: versatile plant blue-light receptors[J]. Trends Plant Sci, 2002, 7(5): 204-210.
[7] CASHMORE A R, JARILLO J A. Cryptochromes: blue light receptors for plants and animals[J]. Science, 1999, 284(5415): 760-765.
[8] LI Q H, YANG H Q. Cryptochrome signaling in plants[J]. Photochem Photobiol, 2007, 83(1): 94-101.
[9] LIU L J, ZHANG Y C. COP1-mediated ubiquitination of CONSTANS is implicated in cryptochrome regulation of flowering inArabidopsis[J]. Plant Cell, 2008, 20(2): 292-306.
[10] CERDAN P D, CHORY J.Regulation of flowering time by light quality[J]. Nature, 2003, 423(6942): 881-885.
[11] JANG S, MARCHAL V. Arabidopsis COP1 shapes the temporal pattern of CO accumulation conferring a photoperiodic flowering response[J]. EMBO J, 2008, 27(8): 1277-1288.
[12] KANG C Y, LIAN H L. Cryptochromes, phytochromes, and COP1 regulate light-controlled stomatal development inArabidopsis[J]. Plant Cell, 2009, 21(9): 2624-2641.
[13] REED J W, NAGPAL P. Mutations in the gene for the red/far-red light receptor phytochrome B alter cell elongation and physiological responses throughoutArabidopsisdevelopment[J]. Plant Cell, 1993, 5(2): 147-157.
[14] MOGLICH A, YANG X. Structure and function of plant photoreceptors[J]. Annu Rev Plant Biol, 2010, 61: 21-47.
[15] CHIU F Y, CHEN Y R. Electrostatic interaction of phytochromobilin synthase and ferredoxin for biosynthesis of phytochrome chromophore[J]. J Biol Chem, 2010, 285(7): 5056-5065.
[16] KOHCHI T, MUKOUGAWA K. The Arabidopsis HY2 gene encodes phytochromobilin synthase, a ferredoxin-dependent biliverdin reductase[J]. Plant Cell, 2001, 13(2): 425-436.
[17] VOSS I, KOELMANN M. Knockout of major leaf ferredoxin reveals new redox-regulatory adaptations inArabidopsisthaliana[J]. Physiol Plant, 2008, 133(3): 584-598.
[18] JEONG S, CLARK S E.Photoperiod regulates flower meristem development inArabidopsisthaliana[J]. Genetics, 2005, 169(2): 907-915.
