
白羊草无性系构件的结构及生长规律
2013-03-14 04:44:14贾娜尔阿汗李海燕杨允菲
草业科学 2013年5期
贾娜尔·阿汗,赵 玉,刘 影,李海燕,杨允菲
(1.新疆自治区普通高校天然产物化学与应用重点实验室 资源与生态研究所 伊犁师范学院化学与生物科学学院,新疆 伊宁 835000; 2.东北师范大学草地科学研究所 植被生态科学教育部重点实验室,吉林 长春 130024)
白羊草(Bothriochloaischaemum)为禾本科孔颖草属,喜温,中旱生植物,耐践踏,适口性好,属优良牧草。广泛分布于新疆境内的低山丘陵地带,垂直分布在海拔480~2 600 m,在伊犁河谷海拔1 000~1 700 m山地地带尤为集中,是伊犁河谷山地草原的建群种,在局部地段形成单优群落[1-2]。目前,国内对白羊草的研究主要集中在种群生态学、群落结构、群落类型划分、抗旱以及抗盐等逆境生理生态特征等方面。山西境内白羊草株丛的生殖枝数和营养枝数比值随株丛高度和丛径均呈二次曲线变化[3],白羊草草地植物量随季节变化呈单峰曲线变化[4],且水热等综合因素对其分布也有影响[5],垂直分布上限可以到海拔1 950 m左右,水平分布范围已由森林草原地带的建群种或优势种扩展到典型草原地带,并从伴生种上升为优势种。徐朗然等[6]和吴东丽等[7]分别从不同的角度对黄土高原区和山西境内的白羊草群落进行了群丛划分。白羊草群落生产力受多种因素的影响,其中水是影响白羊草群落组成和生物量的主导因子[8],并使得群落在季相、结构、生物量等方面发生变化[9]。白羊草具有较强的抗氧化及抵御逆境胁迫的能力[10],在干旱胁迫和复水后野生种的耐旱性强于栽培种[11]。
对无性系的生长和结构进行研究是种群乃至群落动态研究的基础[12],白羊草属于无性系植物,是伊犁河谷山地草原的重要建群种,却鲜见对其无性系特征的研究报道。因此,本研究对伊犁河谷山地草原白羊草无性系构件的结构和生长规律进行初步调查,以期为白羊草的可持续利用和科学管理提供理论依据。
1 研究样地与方法
1.1样地自然概况 研究样地位于伊犁州特克斯县罕盖牧业村(81°19′~82°37′ E,42°22′~42°25′ N),为山地草原类,冬季漫长且较寒冷,夏季不炎热,春温不稳定,秋温下降快,气温年、日较差较大,年均气温5.3 ℃,无霜期118 d,年均降水量383 mm,属温带亚干旱大陆性气候[13]。白羊草为该草地建群种,伴生植物主要有旱雀麦(Bromustectorum)、羊茅(Festucaovina)、芨芨草(Achnatherumsplendens)以及针茅(Stipacapillata)等,该草地主要作为春秋牧场进行刈割利用。
1.2研究方法 于2011年8月20-21日白羊草籽实成熟期,在研究样地内,采用大样本随机取样方法,挖取34个无性系株丛。取样时保持植物地上与地下构件的自然联系和完整,并按无性系分别装袋及标记,带回实验室测定。测定时首先用自来水将根上的泥土冲洗干净,逐一测定分蘖丛的丛径(丛径指分蘖丛的直径,在株丛基部分蘖节部位,按不同方向3次测量的平均值),计数各无性系的生殖分株数和营养分株数,将生殖分株和营养分株从分蘖节处剪下,在自然条件下风干后分别称其生物量。
1.3数据分析 用Microsoft Excel 2003软件对所得数据进行统计分析与作图。用最小值和最大值反映样本的实际范围,各数量指标的平均值反映样本的整体水平,用标准差(SD)反映无性系株丛间的绝对变异度,用变异系数(CV)反映无性系间的相对变异度[12,14]。分析各构件数量及生物量、无性系大小及生物量变化关系的定量刻画模型时,在线性函数、指数函数和幂函数中选择相关性系数最高的函数作为刻画模型[15-17]。
2 结果与分析
2.1无性系构件的数量与结构特征 自然生境中的白羊草无性系一般较均匀地向外扩展,使无性系呈圆形丛生状。因此,可以根据丛径和丛径面积来衡量其无性系的大小。在调查的34个无性系中,白羊草平均丛径为13.5 cm,平均丛径面积为162.1 cm2。丛径的变异系数最小,为32.4%,而生殖分株数量和生物量的变异系数为丛径的近3倍,具有较高的变异。营养分株和总分株数量的变异系数基本相同,且相对较低,表明在各无性系中营养分株和总分株数量是一定的,其差异仅在生物量上体现。调查期,白羊草无性系构件由营养分株和生殖分株组成。在分株数量上,生殖分株平均为19.2个分蘖株,营养分株平均为49.6个分蘖株,营养分株占无性系总分株数量的72.1%,而生殖分株数量仅占27.9%,营养分株数量是生殖分株数量的2.6倍。在生物量分配上,生殖分株生物量占50.6%,营养分株生物量占49.4%,二者的比例为1.02∶1 (表1)。

表1 白羊草无性系构件的数量特征及构件结构(n=34) Table 1 Quantitative characters and structures of the modules in Bothriochloa ischaemum clone (n=34)
2.2无性系构件数量和生物量与分蘖丛大小的关系 经统计分析,在34个白羊草无性系中,白羊草无性系的营养分株数量、生殖分株数量以及总分株数量与丛径均呈显著正相关(P<0.05),并且均以指数的相关性最高(图1);营养分株生物量、生殖分株生物量和分株总生物量也均与丛径呈显著正相关(P<0.05),并且均以指数的相关性最高(图1)。生殖分株生物量与丛径间指数关系的增长率b值,略大于营养分株生物量和分株总生物量与丛径间指数关系的增长率b值。
2.3无性系构件数量和生物量与总分株数量间的关系 在所调查的34个白羊草无性系中,营养分株数量和生物量与总分株数量呈显著正相关(P<0.05),均以幂函数的相关性最高,营养分株与总分株数量无论在数量还是在生物量间均有同样的生长规律。白羊草无性系中生殖分株数量和生物量与总分株数量间均呈显著的线性正相关(P<0.05),表明随着无性系总分株数量的增加生殖分株的数量和生物量具有相同的增长规律。

图1 白羊草无性系不同构件的数量和生物量与无性系大小的观测值及拟合曲线(n=34)Fig.1 Observed data and simulated curves of relationship between both number and biomass of different modules and clonal sizes in B.ischaemum clone (n=34)
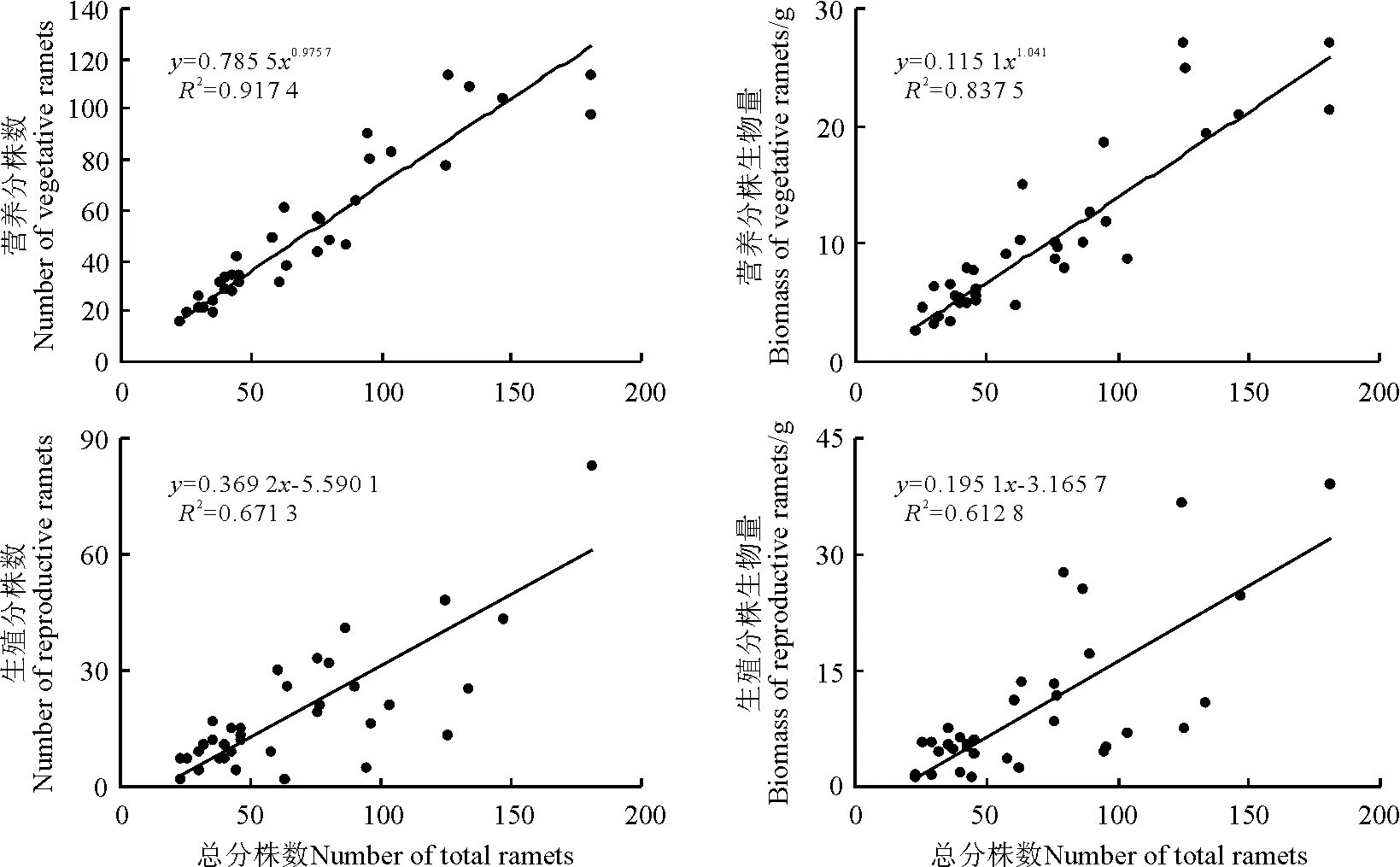
图2 白羊草无性系不同构件的数量和生物量与总分株数量的观测值及拟合曲线(n=34)Fig.2 Observed data and simulated curves of relationship between both number and biomass of different modules and number total ramets in B.ischaemum clone(n=34)
2.4无性系构件数量和生物量与分株总生物量间的关系 经回归分析,白羊草无性系的营养分株数量和生物量与分株总生物量间均呈显著正相关(P<0.05),二者均随着分株总生物量的增加呈幂函数关系增长,且营养分株数量和生物量增长的幂值b值大致相同,表明营养分株数量和生物量与分株总生物量之间具有同速增长规律。
生殖分株数量和生物量与分株总生物量之间均呈显著线性正相关(P<0.05),生殖分株数量和生物量随分株总生物量的增加以相同规律增长(图3)。
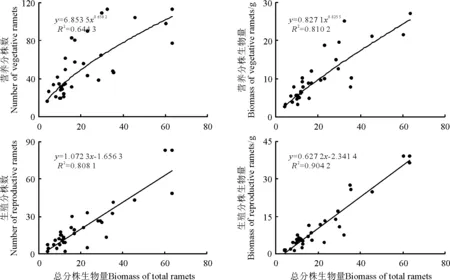
图3 白羊草无性系不同构件的数量和生物量与分株总生物量的观测值及拟合曲线(n=34)Fig.3 Observed data and simulated curves of relationship between both number and biomass of different modules and biomass of total ramets in B.ischaemum clone(n=34)
3 讨论与结论
3.1无性系构件的结构特征 在白羊草籽实成熟期,其无性系构件由营养分株和生殖分株组成,营养分株数量占总分株数量的72.1%,营养分株数量是生殖分株数量的2.6倍,在无性系构件的数量上营养株占绝对优势,表明白羊草处于果后营养期的生长阶段。在生物量分配上,营养分株生物量与生殖分株生物量的比值近1∶1,表明白羊草无性系中营养分株和生殖分株的生物量几乎相等。这与李红[14]等研究野大麦(Hordeumbrevisubulatum)无性系构件的能量分配机制相同,白羊草无性系也对营养生长和生殖生长的能量投入大体均等,通过产生小而数量多的营养分株和大而数量少的生殖分株来调节能量分配的均衡。这一特征是白羊草无性系生存与发展所适应的机制之一。
3.2无性系构件与无性系大小的关系 杨允菲等[13]对松嫩平原贝加尔针茅无性系构件的结构及其生长规律的研究中曾指出,指数函数可以用于描述构件间由缓速到加速的异速增长过程,幂函数关系则可以视为由加速到缓速的异速增长过程,而直线函数关系则是同速增长过程。由此说明,在白羊草无性系的扩展过程中,各构件无论在数量上,还是在生物量上均呈现为从缓速到加速的异速增长规律,即随着白羊草无性系丛径的增长,无性系的营养分株数量和生物量、生殖分株数量和生物量及总分株数量和分株总生物量均以指数关系的变化规律生长。白羊草无性系中生殖分株生物量随丛径增长的速度略大于营养分株生物量和分株总生物量随丛径增长的速度。
3.3无性系构件与总分株数量、分株总生物量间的关系 营养分株数量和生物量与总分株数量及总生物量之间均呈幂函数关系,且均呈现出由加速到缓速的异速增长规律。而生殖分株数量和生物量与总分株数量及分株总生物量之间均呈线性相关关系,且均具始终不变的同速增长规律。以上对白羊草无性系各构件结构与生长规律所作的结论在其它生长期是否发生变化有待进一步研究。
[1] 新疆植物志编辑委员会.新疆植物志(第6卷)[M].乌鲁木齐:新疆科技卫生出版社,1996.
[2] 米吉提.胡达拜尔地,徐建国.新疆高等植物检索表[M].乌鲁木齐:新疆大学出版社,2000:672-673.
[3] 董宽虎,米佳.白羊草种群繁殖的数量特征[J].草地学报,2006,14(3):210-213.
[4] 黄锋华,董宽虎.白羊草灌丛草地植物量及优势种牧草营养动态研究[J].草原与草坪,2007(2):14-17.
[5] 程杰,呼天明,程积民.黄土高原白羊草种群分布格局对水热梯度的响应[J].草地学报,2010,18(2):167-171.
[6] 徐朗然,张继敏,丁士友.黄土高原白羊草草原的基本特征及其地理学意义[J].西北植物学报,1997,17(1):88-93.
[7] 吴东丽,上官铁梁,高洪文.山西东南部白羊草群落植物种多样性研究[J].草地学报,2002,10(4):237-243.
[8] 李琪,张金屯,高洪文.山西高原三种白羊草群落的生物量研究[J].草业学报,2003,12(1):53-58.
[9] 张娜,梁一民.干旱气候对白羊草群落土壤水分和地上部生长的初步观察[J].生态学报,2000,20(6):964-970.
[10] 单长卷,韩蕊莲,梁宗锁.干旱胁迫下黄土高原4种乡土禾草抗氧化特性[J].生态学报,2012,32(4):1174-1184.
[11] 刘娟,董宽虎.干旱胁迫及复水处理对白羊草抗旱生理特性的影响[J].草原与草坪2011,31(2):74-78.
[12] 杨允菲,张宝田.松嫩平原贝加尔针茅无性系构件的结构及生长规律[J].应用生态学报,2005,16(12):2311-2315.
[13] 新疆维吾尔自治区地方志委员会.新疆维吾尔自治区地方志丛书·特克斯县志[M].乌鲁木齐:新疆人民出版社,2004.
[14] 李红,杨允菲,包国章.两种生境中虎尾草无性系分蘖株构件的定量研究[J].东北师大学报(自然科学版),2002,34(4):80-86.
[15] 杨允菲,张宝田.松嫩平原不同生境条件下羊草无性系的生长规律[J].应用生态学报,2006,17(8):1419- 1423.
[16] 赵玉,贾娜尔,李海燕,等.新疆伊犁河谷平原狗牙根无性系构件的生长[J].应用生态学报,2009,20(4):779- 784.
[17] 宋金枝,李海燕,周丽威.长白山区华北翦股颖无性系构件的结构及生长[J].生态学杂志,2011,30(10):2145-2148.
猜你喜欢
小哥白尼(野生动物)(2021年3期)2021-07-21 02:28:34
中国生殖健康(2020年8期)2021-01-18 03:05:08
中国生殖健康(2020年2期)2021-01-18 02:51:10
今日农业(2020年19期)2020-12-14 14:16:52
中国生殖健康(2018年1期)2018-11-06 07:14:48
中国生殖健康(2018年2期)2018-11-06 07:10:32
小学阅读指南·低年级版(2017年4期)2017-04-24 13:33:52
中国科技信息(2015年2期)2015-11-16 08:18:32
植物营养与肥料学报(2014年1期)2014-03-11 20:24:38
中国烟草学报(2012年3期)2012-04-10 12:50:56
