
纤维母细胞生长因子10基因在角膜混浊小鼠中的克隆测序与表达研究*
2013-03-04 05:12吴刘成张柳柳王胜洁倪尧尉邵义祥
重庆医学 2013年36期
吴刘成,张柳柳,李 瑶,王胜洁,季 韧,倪尧尉,邵义祥△
(1.南通大学比较医学研究所,江苏南通226001;2.南通大学医学院,江苏南通226001)
在哺乳动物正常发育过程中,眼睑生长覆盖眼睛、融合,而后重新开放。小鼠在正常发育过程中,胚胎期约11d开始出现原始的眼睑即单层细胞上皮,随着眼睑不断生长发育覆盖了角膜,约第16.5d眼睑上缘和下缘融合,完成胚胎期眼睑闭合,出生后约14d眼睑重新开放[1-2]。B6-Co(C57BL/6-corneal opacity)小鼠是通过乙基亚硝基脲(ENU)诱变技术获得并建立的出生时眼睑开放(EOB)突变系小鼠,出生后角膜逐渐混浊,角膜混浊发生率约为43.1%[3],突变基因定位于13号染色体上60cM附近[4]。纤维细胞生长因子10(fibroblast growth factor 10,Fgf10)基因突变小鼠也表现为出生时眼睑开放[5],并且该基因位于突变基因定位区间。因此,本实验对该基因进行克隆测序分析,并利用实时荧光聚合酶链式反应(PCR)技术检测Fgf10基因的表达,探讨Fgf10基因与B6-Co小鼠EOB表型的关联,为研究人类遗传性眼睑缺陷形成机制提供理论依据。
1 材料与方法
1.1 实验动物 本实验所用B6和B6-Co小鼠由南通大学实验动物中心提供,小鼠饲养在清洁级屏障动物房内,实验动物生产许可证号:SCXK(苏)2008-0010,使用许可证号:SYXK(苏)2012-0030。用B6与B6-Co小鼠交配,次日清晨检栓,妊娠16.5d,麻醉取胚胎。
1.2 主要试剂与仪器 Trizol、琼脂糖为加拿大BBI公司生产;逆转录试剂盒(RT)、Taq DNA Polymerase为加拿大Fermentas公司生产;引物合成、UNIQ-10柱式DNA胶回收试剂盒为生工生物(上海)股份有限公司生产;T载体试剂盒为美国Promega公司生产;TOP10感受态细胞、质粒小抽试剂盒为南京天根公司生产;EcoRⅠ为大连宝生物工程有限公司生产;荧光染料试剂盒为美国Biotium公司生产。超净工作台为苏净集团安泰公司生产;Centifuge 5417R冷冻离心机、Eppendorf AG 22331Hamburg PCR仪为德国Eppendorf公司生产;DYY-6B型稳压稳流电泳仪为北京市六一仪器厂生产;凝胶成像系统为江苏省捷达科技发展有限公司生产;Step One Real-Time PCR System为美国ABI公司生产。
1.3 方法
1.3.1 引物设计 在Gene Bank中查找并下载Fgf10mRNA序列,以mRNA为模板设计部分引物,引物之间有重叠区域,Fgf10-a正向引物:5′-CAC CGT CCT GTC ATC TGT TT-3′,反向引物:5-TTC TTC TAT GTT TGG ATC GTC A-3′;Fgf10-3正向引物:5′-ATG GAA TCA AGT ATT ATC ACC-3′,反向引物:5′-AGT GTT TGC CAA GGT TTT-3′;Fgf10-c正向引物:5′-GCA CAA CCA AAG GAG C-3′,反向引物:5′-GAT CTA CAG AGC GAG GG-3′;Fgf10-d正向引物:5′-GCA CAA CCA AAG GAG C-3′,反 向引物:5′-GAT CTA CAG AGC GAG GG-3′;Fgf10-e正向引物:5′-GCA CAA CCA AAG GAG C-3′,反向引物:5′-GAT CTA CAG AGC GAG GG-3′。设计实时荧光PCR引物,引物至少跨2个外显子,Fgf10正向引物:5′-TCC GCT GGA GAA GGC TGT-3′,反向引物:5′-CAA CTC CGA TTT CCA CTG ATG T-3′;甘油醛-3-磷酸脱氢酶(GAPDH)为内参,正向引物:5′-GGA GCG AGA CCC CAC TAA C-3′,反向引物:5′-GGC GGA GAT GAT GAC CCT-3′。
1.3.2 组织总RNA的提取和逆转录 剪取胚胎16.5dB6和B6-Co小鼠皮肤组织,放入匀浆器中,置于冰上,加入1mL TRizol,充分匀浆,按照试剂盒说明操作,最后用焦碳酸二乙(DEPC)处理水溶解获得RNA,核酸定量仪测定RNA浓度;按照RT试剂盒说明将RNA逆转录合成cDNA。
1.3.3 基因测序 PCR反应体系,10×PCR缓冲液 (buffer,含 Mg2+)5.0μL,dNTP(10mmol/L)1μL,上游引物(50 μmol/L)0.5μL,下游引物(50μmol/L)0.5μL,模板cDNA 2.5ng,Taq DNA polymease(5U/μL)0.25μL,补灭菌去离子水至50μL。扩增条件,94℃5min;94℃1min,63℃30s;72℃30s,35个循环,72℃5min,根据引物熔解温度(Tm)值和扩增长度调整退火温度、变性时间及延伸时间。PCR产物经1%琼脂糖凝胶电泳检测,成像保存。PCR产物的纯化(胶回收),方法参照胶回收试剂盒进行。基因测序、回收产物与相应引物寄送上海生工生物股份有限公司测序。
1.3.4 基因的克隆测序 将发现突变的纯化PCR产物与T载体连接,2×buffer 5μL,目的DNA 1~3μL,T载体1μL,T4DNA Ligase 1μL,补灭菌去离子水至10μL,4℃ 过夜。取5μL链接产物和50~100μL感受态细胞加入预冷的离心管中;冰浴5min;42℃热休克90s;冰浴30min;加入200μL Luria-Bertani(LB)培养液,37℃,250r/min,振荡培养45min;取120~200μL菌液涂于1.5%琼脂LB平板(添加氨苄青霉素、β-半乳糖苷酶和异丙基硫代半乳糖苷);37℃培养过夜;挑取白斑,扩培摇菌12~14h;提取质粒DNA分子;电泳检测;EcoRⅠ酶切释放目的片段,电泳检测,寄送上海生工生物股份有限公司测序。
1.3.5 实时荧光PCR 以GAPDH基因为内参,每个样品设置3个重复,反应体系为荧光染料预混液10μL,正、反引物各1μL,cDNA 1μL,补灭菌去离子水至20μL。反应条件,95℃4min;95℃20s,60℃20s,72℃30s并读取荧光值,45次循环,循环后设置55~90℃每隔0.3℃读荧光值生成熔解曲线。所有设定保存后运行程序。反应完成后,用相对定量分析方法分析各样品表达量。
1.4 统计学处理 PCR数据采用Stata7.0软件分析,计量资料用±s表示,组间比较采用t检验,检验水准α=0.05,以P<0.05为差异有统计学意义。
2 结 果
2.1 Fgf10基因测序及克隆测序结果 分别以表1中前5对引物对角膜混浊小鼠的基因进行扩增,从左向右依次电泳,得到大小不同的5个基因片段(图1A)。对以上5个片段割胶回收后分别进行测序,发现第4个片段基因有突变(图1B)。将第4个基因片段用T载体连接,对该基因片段进行克隆。通过蓝白斑筛选出重组质粒、质粒的鉴定,从上到下分别是开环质粒,环状质粒和超螺旋质粒,胶回收中间条带的环状条带(图1C)。重组质粒的EcoRⅠ酶切片段电泳图。限制性内切酶作用于重组质粒后,将环状重组质粒切成2个大小不同的片段,胶回收目的基因(图1D)。分离出目的基因,再对目的基因测序分析,并通过数据库比对发现在该基因第3外显子1 914bp和1 915bp之间插入碱基A(图1E)。

图1 Fgf10基因克隆测序流程图
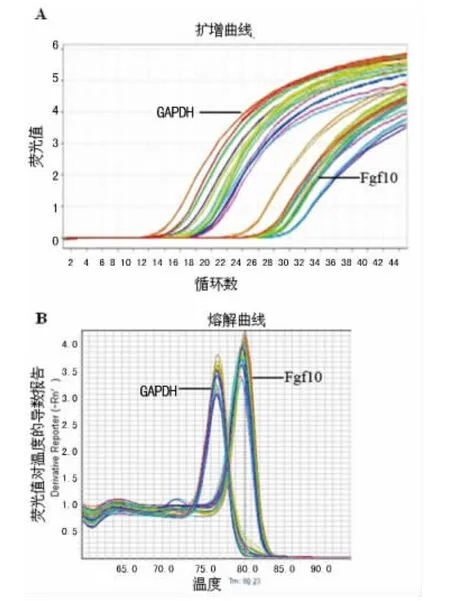
图2 Fgf10基因扩增曲线和熔解曲线图

图3 实时荧光PCR检测Fgf10基因在B6-Co小鼠中的表达
2.2 荧光定量PCR分析结果 由图2可知,GAPDH、Fgf10 2个基因的扩增曲线均接近“S”型曲线,基因扩增效率良好;熔解曲线分析发现2个基因的PCR产物均呈较为锐利的单峰,无杂峰。Fgf10基因mRNA表达水平在B6-Co小鼠中明显低于B6小鼠,差异有统计学意义(P<0.05),见图3。
3 讨 论
眼睑是眼的门户,能够保护眼球避免受外伤与强烈光线刺激,协助瞳孔调节进入眼内光线,起到维持眼位及眼功能的作用;眼睑还与其他眼附属器一起共同发挥功能作用,保护角膜免受微生物感染。因此,眼睑发育缺陷将使眼的门户受损,调节和保护功能受限或缺失,导致各种眼部疾病[6-7]。人类遗传性眼睑缺陷并伴有其他眼部疾病是一种遗传性/先天性疾病,将有可能进一步造成严重的眼科疾病,如角膜炎、角膜感染、角膜混浊,重则致盲,现在临床多采用手术植皮的方法进行眼睑的修复,没有对应的动物模型可供研究。如何做到早期的诊断、预防和治疗,必须首先建立起研究技术平台,进行基础理论和实验技术的研究,建立可利用的动物模型深入进行研究是最佳途径。
小鼠胚胎眼睑闭合,是一个包括细胞增殖和迁移,细胞形态,牵涉许多生理和病理过程的动态发育过程[8]。绝大多数的哺乳动物的眼睑发育都要经历由融合至再分离的过程,并且这个过程能以相对恒定的程序发生。眼睑覆盖在眼球的前方,起保护眼球的作用,新生小鼠正常情况下眼睑闭合,若眼睑不能闭合,可能由于新生鼠抵抗力低,受外界病原体感染及异物刺激,引起角膜感染及角膜混浊。B6-Co小鼠眼睑发育缺陷导致角膜发育异常和先天性眼表疾病,该小鼠角膜混浊的病理过程呈典型的炎性反应,与人类角膜炎的病理变化过程相似[9],值得进一步研究开发其价值。
Fgf10缺失小鼠在出生时表现出眼睑开放并伴有多器官发育缺陷[10]。Jin等[11]已经证实Fgf10通过调节表皮生长因子受体(EGFR)信号通路来促进细胞增殖和迁移以促进眼睑的发育。还有研究表明,Fgf10的功能可能是调节细胞骨架的活性和基因转录,在小鼠眼睑发育中,可能与Wnt信号通路调节有关[12]。通过分离和培养野生型和Fgf10缺陷型胚胎眼睑的角质形成细胞,行体外划痕实验,证明小鼠眼睑上皮细胞的增殖、迁移能力受到严重阻碍,从而导致新生小鼠眼睑上下缘周皮的完整性存在缺陷,Fgf10信号中断也使得激活素βB和Tgfa下调,且延迟了声音刺猬(Shh)蛋白的表达[13]。
有研究表明,基因的编码区或非编码区发生点突变后,可以使机体产生某种疾病[14-15]。本实验对该基因进行了PCR扩增,通过克隆测序发现在Fgf10基因第3外显子1 914bp和1 915bp之间插入碱基A(图1),位于3′末端非编码区。为进一步了解该突变基因在B6-Co小鼠中的作用,采用实时荧光PCR技术检测Fgf10基因的表达变化,结果表明该基因在B6-Co小鼠胚胎E 16.5d的表达明显下调(P<0.05)(图3),提示该突变位点可能影响了其基因的表达,或直接影响了功能蛋白质的翻译与剪接,是否B6-Co小鼠中存在其他突变基因而影响了Fgf10基因的表达调控也值得深入探讨。作用于基因表达调控的因素很多,其具体机制需进一步深入研究,从而为人类眼科类疾病的诊断、治疗及预防提供参考依据。
[1]Ohuchi H.Wakayama symposium:epithelial-mesenchymal interactions in eyelid development[J].Ocul Surf,2012,10(4):212-216.
[2]Harris MJ,Juriloff DM.Eyelid development and fusion induced by cortisone treatment in mutant,lidgap-Miller,fetal mice.A scanning electron microscope study[J].Embryol Exp Morphol,1986(91):1-18.
[3]邵义祥,吴宝金,薛整风,等.遗传性角膜基质变性小鼠及其突变基因的定位[J].南京师大学报,2006,29(2):99-102.-
[4]蒋荧梅,刘春,吴刘成,等.SNP标记对角膜混浊小鼠突变相关基因的精细定位[J].遗传,2010,32(5):486-491.
[5]Huang J,Dattilo LK,Rajagopal R,et al.FGF-regulated BMP signaling is required for eyelid closure and to specify conjunctival epithelial cell fate[J].Development,2009,136(10):1741-1750.
[6]Qu X,Pan Y,Carbe C,et al.Glycosaminoglycan-dependent restriction of FGF diffusion is necessary for lacrimal gland development[J].Development,2012,139(15):2730-2739.
[7]Puk O,Esposito I,Söker T,et al.A new Fgf10mutation in the mouse leads to atrophy of the harderian gland and slit-eye phenotype in heterozygotes:a novel model for dryeye disease?[J].Invest Ophthalmol Vis Sci,2009,50(9):4311-4318.
[8]Herr DR,Lee CW,Wang W,et al.Sphingosine 1-Phosphate Receptors Are Essential Mediators of Eyelid Closure during Embryonic Development[J].J Biol Chem,2013,288(41):29882-29889.
[9]刘春,袁志云,杨菲菲,等.C57BL/6小鼠葡萄球菌性角膜感染模型的建立及相关研究[J].眼科研究,2010,28(3):199-202.
[10]Tsau C,Ito M,Gromova A,et al.Barx2and Fgf10regulate ocular glands branching morphogenesis by controlling extracellular matrix remodeling[J].Development,2011,138(15):3307-3317.
[11]Jin C,Yin F,Lin M,et al.GPR48regulates epithelial cell proliferation and migration by activating EGFR during eyelid development[J].Invest Ophthalmol Vis Sci,2008,49(10):4245-4253.
[12]Jin C,Chen J,Meng Q,et al.Deciphering gene expression program of MAP3K1in mouse eyelid morphogenesis[J].Dev Biol,2013,374(1):96-107.
[13]Wu CI,Hoffman JA,Shy BR,et al.Function of Wnt/βcatenin in counteracting Tcf3repression through the Tcf3-β-catenin interaction[J].Development,2012,139(12):2118-2129.
[14]吴伟,胡锦林,鞠海星,等.散发性大肠癌K-ras基因点突变研究及其临床病理意义[J].医学研究杂志,2010,39(12):53-57.
[15]夏海龙,陈晓文,陈歆,等.弥漫性大B细胞淋巴瘤PRDM1基因5′非编码区突变的研究[J].安徽医科大学学报,2010,45(1):27-30.
猜你喜欢
保健医苑(2020年8期)2020-08-19
食品科学(2018年10期)2018-05-23
中国医药指南(2017年3期)2017-11-13
西南医科大学学报(2015年1期)2015-08-22
中国医疗美容(2015年2期)2015-07-19
中国医疗美容(2015年2期)2015-07-19
中国医疗美容(2015年2期)2015-07-19
中国医疗美容(2015年1期)2015-07-12
医学研究杂志(2015年8期)2015-06-22
医学研究杂志(2015年12期)2015-06-10
