
长白山地区熊蜂的访花偏爱性研究
2012-12-26 08:59任炳忠尚利娜
东北师大学报(自然科学版) 2012年1期
任炳忠,尚利娜,陈 新,韩 叶,徐 燕
(东北师范大学生命科学学院,吉林 长春 130024)
长白山地区熊蜂的访花偏爱性研究
任炳忠,尚利娜,陈 新,韩 叶,徐 燕
(东北师范大学生命科学学院,吉林 长春 130024)
对长白山地区7种熊蜂访问的主要蜜源植物的花形、花色、花味及花粉的形态结构进行了研究,总结出了熊蜂偏爱访问的蜜源植物的花部特征.结果表明:熊蜂偏爱访问具有圆锥花序、总状花序和聚伞花序的花,对于单生花,通常花形较大;访问的花冠类型以辐射对称和两侧对称为主;花多具有蜜腺,分泌花蜜;色泽艳丽,常为黄色或蓝紫色;花粉粒较大,形状多为长球形,具三孔沟,表面具网状纹饰;花的气味成分主要为酸类和酯类化合物,其中含有较高浓度的羟基丙酮、乙酸、甲酸、苯酚和9,12-十八碳二烯酸乙酯.
长白山地区;熊蜂;访花偏爱性
由于移动能力的限制以及繁衍子代的需要,植物与昆虫之间发生了极为密切的联系[1-6],其中所产生的最为特化的结果是植物演化出虫媒花,昆虫演化出传粉昆虫[6].在自然界中,传粉昆虫的种类繁多,其中膜翅目蜜蜂科昆虫的种类最多,数量最大;同时蜜蜂科独特的形态和生理结构以及社会性群体生物学特性,使其成为绝大多数农作物的最佳授粉者,而其中的熊蜂更是温室作物授粉中的佼佼者.除能对野生植物授粉以外,熊蜂已经广泛应用于人工栽培的番茄、辣椒、茄子、甜瓜、西瓜、黄瓜、南瓜、草莓、树莓、黑莓、蓝莓、醋栗、酸果蔓、苹果、桃、樱桃、猕猴桃、杏、李等作物和水果的授粉过程[7].近年来,熊蜂授粉已经实现了产业化和商品化,荷兰、比利时、英国、以色列、新西兰、土耳其、美国、加拿大等国相继建立了工厂化周年繁育和出售熊蜂的专业公司[8-15].20世纪90年代初,中国农业科学研究院蜜蜂研究所首次成功人工繁育了熊蜂种群,开创了我国熊蜂人工繁育的先河.到目前为止,我国已从本土十几种熊蜂中,筛选出了3种具有重要授粉价值的良种熊蜂,即明亮熊蜂(Bombuslucorum(L.))、红光熊蜂(BombusignitusSmith)和火红熊蜂(BombuspyrosomeMorawitz),它们为温室西红柿、青椒、茄子、黄瓜、冬瓜等蔬菜和桃、油桃等果树授粉效果显著[16-22].
目前,对传粉昆虫的研究已不局限于简单地描述传粉者种类,更深入的研究主要包括两个方面,即传粉昆虫访花行为的研究和传粉昆虫访花偏爱性的研究,这两个方面直接决定了昆虫传粉的成功率.同时,环境温度、风速、光照强度、降雨等各种非生物因子同样影响传粉昆虫的访花行为.传粉昆虫为植物传粉是为了觅食富含营养的花粉、花蜜或其他特殊的回报[23-24].传粉昆虫对花朵的认识主要有花朵构成和花朵形态两个方面[25],它们也影响着传粉昆虫的传粉效率.花朵构成是指花的颜色、花朵大小、气味和给访花者提供的回报等所有单花特征;花朵形态是指花在某一时刻的开放程度和花序上的排列.长白山地区熊蜂及蜜源植物种类丰富,本文以该地区7种主要的熊蜂,即密林熊蜂(BombuspatagiatusNylander)、明亮熊蜂(B.lucorum)、红光熊蜂(B.ignitus)、小峰熊蜂(BombushypocritePerez)、眠熊蜂(Bombushypnorum(L.))、谦 熊 蜂 (BombusmodestusEversmann)和关熊蜂 (BombusconsobrinusDahlbom)为对象,调查了熊蜂访问的主要蜜源植物的种类,探讨了主要蜜源植物的花朵构成与熊蜂访花行为之间的关系,旨在明确熊蜂的访花偏爱性,扩大熊蜂授粉的蜜源植物谱,寻求对熊蜂具有引诱作用的物质,为熊蜂的授粉应用及开发新的传粉昆虫引诱剂提供基础材料.
1 研究方法
1.1 主要蜜源植物花的形态结构及颜色
2007—2009年的6—9月,选择晴朗天气,记录长白山地区7种熊蜂访问的主要蜜源植物的种类;统计和描述主要蜜源植物花的结构、颜色等花朵构成特征,分析这些特征与熊蜂访花行为之间的关系.[26]
1.2 主要蜜源植物花的气味
2007—2009年的6—9月,采集吉林省长白山地区熊蜂访问的6种主要蜜源植物,即珍珠梅(Sorbariakirilowii(Reqel)Maxim.)、蚊子草 (Filipendulapalmata(Pall.)Maxim.)、高 山 芹(Coelopleurumsaxatile(Turcz.)Drude)、大 独 活 (AngelicagigasNakai)、柳 兰 (Chamaenerion angustifolium(L.)Scop.)和白车轴草(TrifoliumrepensL.)处于盛开期的鲜花各1kg,在常温、常压下,分别用乙醇浸泡3次,每次12h,合并滤液后用旋转蒸发仪减压浓缩,获得6种蜜源植物花的乙醇提取物.
利用气相色谱-质谱联用仪(Agilent 5975)对6种植物花的乙醇提取物成分进行分析.实验条件如下:气相色谱载气为He,进样量为1μL,色谱柱为HP-Wax,60m×0.25mm×0.25μm,汽化室温度为250℃,初始温度为50℃,时间为3min,升温速率为10℃/min,终温为250℃,保持90min,连接杆温度为250℃,质谱电离方式为EI,电子能量为70eV,离子源温度为230℃.化合物鉴定利用计算机谱库检索程序得出.通过数据处理系统检索与标准谱图进行对照、复合,再结合有关文献进行人工谱图解析,确认各个化学成分.并通过微机数据处理系统,按峰面积归一化法进行定量分析,求得各化学成分的相对含量.[27]
1.3 主要蜜源植物的花粉形态结构
2007—2009年的6—9月,采集吉林省长白山地区熊蜂访问的8种主要蜜源植物,即广布野豌豆(ViciacraccaL.)、大山黧豆(CathyrlsdavidiiHance)、珍珠梅(Sorbariakirilowii(Reqel)Maxim.)、蚊子草(Filipendulapalmata(Pall.)Maxim.)、柳叶绣线菊(SpiraeasalicifoliaL.)、白车轴草(TrifoliumrepensL.)、大独活(AngelicagigasNakai)和升麻(AruncussylvesterKostel.),分别选取成熟花粉,经干燥、喷金,置于扫描电子显微镜(HITACHI S-570)中观察和拍照.观察花粉的极面、赤道面、外壁纹饰等,并测量花粉的大小.每次测量以20个花粉为标准,取平均值.
2 结果与分析
2.1 主要蜜源植物花的形态结构及颜色
通过野外观察和记录,长白山地区熊蜂访问的主要蜜源植物有草地风毛菊(Saussureaamara(L.)DC.)、万寿菊(TageteserectaL.)、对叶菊(ZinniaeIegansL.)、兴安乳菀(GalatelladahuricaDC.)、假升 麻 (AruncussylvesterKostel.)、珍 珠 梅 (Sorbariakirilowii(Reqel) Maxim.)、蚊 子 草(Filipendulapalmata(Pall.)Maxim.)、山 刺 玫 (RosadavuricaPall.)、柳 叶 绣 线 菊 (Spiraea SalicifoliaL.)、高山芹(Coelopleurumsaxatile(Turcz.)Drude)、东北羊角芹(AegopodiumalpestreLedeb)、大 独 活 (AngelicagigasNakai)、黑 水 当 归 (AngelicaamurensisSchischk.)、白 车 轴 草(TrifoliumrepensL.)、大 山 黧 豆 (CathyrlsdavidiiHabnce)、多 茎 野 豌 豆 (ViciamulticaulisLedeb.)、广布野豌豆(ViciacraccaLinn.)、歪头菜(ViciaunijugaA.B.)、胡枝子(LespedezabicolorTurcz.)、野小豆(PhaseolusminimusRoxb.)、类叶升麻(ActaeaasiaticaHara)、高山乌头(Aconitum monanthumNakai)、两色乌头(AconitumalboviolaceumKom.)、深山唐松草(ThalictrumtuberiferumMaxim.)、朝鲜铁线莲(ClematiskoreanaKom.)、西伯利亚铁线莲(Clematissibirica(L.)Mill.)、柳兰(Chamaenerionangustifolium(L.)Scop.)、白花败 酱 (PatriniavillosaJuss.)、华水苏 (Stachys chinensisBunge ex Benth.)、益母草(LeonurusjaponicusHoutt.)、毛蕊老鹳草(GeraniumeriostemonFisch.ex DC.)、接骨木(SambucuswillamsiiHance)、朝鲜落新妇(AstilbekoreanaNakai)等.以上蜜源植物分别隶属于豆科、菊科、蔷薇科和伞形科等.
从熊蜂访问的蜜源植物的种类可以看出,熊蜂对蜜源植物具有明显的选择性.熊蜂偏爱访问具有圆锥花序、总状花序和聚伞花序的花朵.对于单生花,通常花均较大,能够承受熊蜂的重量.熊蜂访问的花冠类型以辐射对称、两侧对称为主,以便于熊蜂着落.花多具有蜜腺,可分泌花蜜,作为熊蜂访花的回报.从花的颜色来看,熊蜂访问的花色泽艳丽,常为黄色或蓝紫色.
2.2 主要蜜源植物花的气味
本研究对熊蜂访问的6种主要蜜源植物花的乙醇提取物成分进行了分析,化合物按保留时间由小到大顺序排列,结果见表1.从已鉴定出的成分来看,6种植物鲜花乙醇提取物成分主要为酸类、酯类、酮类化合物,此外还含有醇类、酚类化合物.在蚊子草(F.palmate)鲜花乙醇提取物中,乙酸、9,12-十八碳二烯酸乙酯和n-十六烷酸含量较高;在珍珠梅(S.sorbifolia)鲜花乙醇提取物中,乙酸、苯酚和羟基丙酮含量较高;在高山芹(C.saxatile)鲜花乙醇提取物中,乙酸、氢醌和n-十六烷酸含量较高;在大独活(A.gigas)鲜花乙醇提取物中,乙基-α-d-吡喃葡萄糖苷含量最高,其次为乙酸和羟基丙酮;在柳兰(C.angustifolium)鲜花乙醇提取物中,n-十六烷酸、5-(羟甲基)-2-呋喃甲醛、乙酸的含量最高;在白车轴草(T.repens)鲜花乙醇提取物中,乙酸、n-十六烷酸、棕榈酸(十六烷酸)乙酯、9,12-十八碳二烯酸乙酯的含量较高.对6种植物鲜花乙醇提取物的成分进行比较分析可以看出,6种植物鲜花乙醇提取物中均含有羟基丙酮、乙酸、甲酸、苯酚和9,12-十八碳二烯酸乙酯,且这些化合物在6种植物中的含量均较高.

表1 长白山地区6种蜜源植物花乙醇提取物的化学成分 %
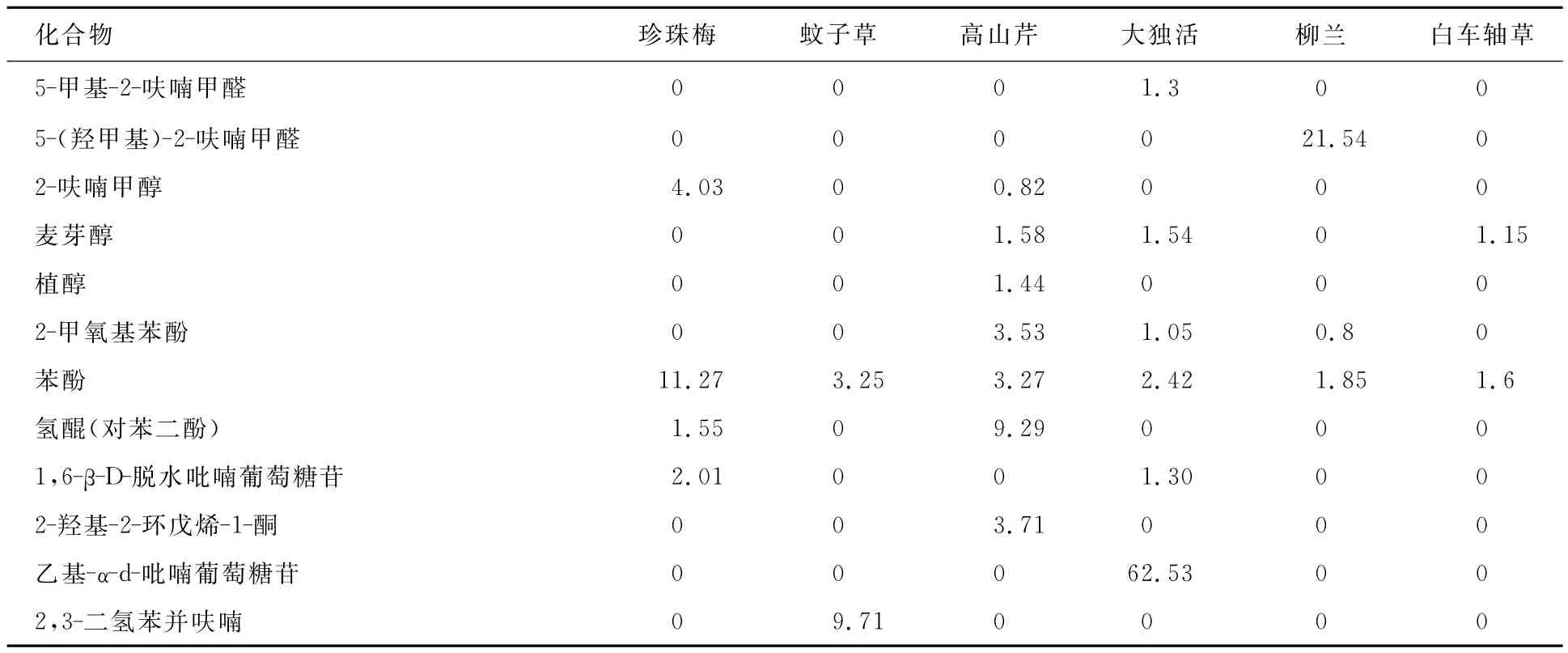
续表1 %
2.3 主要蜜源植物花粉的形态结构
对长白山地区熊蜂访问的8种主要蜜源植物的花粉形态结构进行了观察,结果如下.
(1)广布野豌豆(ViciacraccaL.).花粉长球形,极面观圆形,赤道面窄椭圆形,极轴长36.7(32.3~38.4)μm,赤道轴长21.6(19.2~22.9)μm,极轴与赤道轴之比(P/E)为1.69(1.67~1.77).具三孔沟,沟缘较厚,略呈微波形,孔膜高升.表面具细网状饰纹,网脊较矮(见图1中1A、1B).
(2)大山藜豆(CathyrlsdavidiiHance).花粉长球形,极面观三裂圆形,赤道面窄椭圆形,极轴长34.6(32.5~37.4)μm,赤道轴长25.2(23.9~26.4)μm,极轴与赤道轴之比为1.37(1.23~1.56).具三孔沟,沟细长、边缘整齐.表面具网状饰纹,网眼形状不规则(见图1中2A、2B).
(3)珍珠梅(Sorbariakirilowii(Reqel)Maxim.).花粉长球形,极面观三裂圆形,赤道面观长椭圆形,极轴长24.6(22.3~26.0)μm,赤道轴长14.1(1.52~2.06)μm,极轴与赤道轴之比为1.74(1.52~2.06),具三孔沟,沟狭长,表面具条纹状饰纹(见图1中3A、3B).
(4)蚊子草(Filipendulapalmata(Pall.)Maxim.).花粉长球形,极面观三裂圆形,赤道面窄椭圆形,极轴长25.5(24.4~26.7)μm,赤道轴长15.2(13.1~16.9)μm,极轴与赤道轴之比为1.68(1.58~1.98),具三孔沟,表面具颗粒状纹饰(见图1中4A、4B).
(5)柳叶绣线菊(SpiraeasalicifoliaL.).花粉长球形,极面观三裂圆形,赤道面窄椭圆形,极轴长17.8(12.3~20.8)μm,赤道轴长12.3(9.7~16.1)μm,极轴与赤道轴之比为1.45(1.27~1.9),具三孔沟,表面具网状饰纹(见图1中5A、5B).
(6)白车轴草(TrifoliumrepensL.).花粉长球形,极面观三裂圆形、平坦,赤道面窄椭圆形,极轴长34.1(31.2~36.5)μm,赤道轴长18.3(17.6~19.1)μm,极轴与赤道轴之比为1.86(1.77~1.98),具三孔沟,沟长裂至两极,孔膜不突出或突出不明显,表面具不明显的网状纹饰(见图1中6A、6B).
(7)大独活(AngelicagigasNakai).花粉超长球形或长球形,极面观三裂圆形,赤道面窄椭圆形,极轴长26.6(24.9~29.4)μm,赤道轴长13.0(11.9~15.4)μm,极轴与赤道轴之比为2.05(1.91~2.19),具三孔沟,沟膜具较大的颗粒,表面具网状纹饰(见图1中7A、7B).
(8)升麻(AruncussylvesterKostel.).花粉粒长球形,极面观三裂圆形,赤道面扁圆形或圆形,极轴长27.3(25.9~28.0)μm,赤道轴长17.5(13.6~19.6)μm,极轴与赤道轴之比为1.56(1.41~1.90),具三孔沟,表面具颗粒状纹饰(见图1中8A、8B).
形态指数(P/E)反映了花粉的形状.形态指数越趋近于1,说明花粉越趋向于球形;比1越大,说明花粉形状越长.8种虫媒花P/E平均值为1.68±0.41,表明花粉粒的形态多为长球形.Erdtman把花粉的大小划分为6个等级(以花粉最长轴的长度来表示):小于10μm的为非常小孢粉,10~25μm的为小孢粉,25~50μm的为中等孢粉,50~100μm的为大孢粉,100~200μm的为非常大孢粉,大于200μm的为巨孢粉[28].根据此标准对花粉大小进行分级,8种蜜源植物的花粉为小孢粉和中等孢粉.

图1 长白山地区熊蜂访问的8种蜜源植物的花粉形态(A赤道面观,B极面观)
熊蜂访问的蜜源植物的花粉形状主要为长球形,多为中等孢粉,萌发孔的类型为三孔沟,外壁纹饰多为深刻而清晰的网状纹饰,网状雕纹的网眼中往往还有微小的颗粒状次级雕纹.
3 讨论
当传粉昆虫对花朵进行选择时,会表现出对某种特定类型的花朵的连续采集,这种对花朵的选择行为被称为“访花偏爱性”[29].2000多年前,亚里士多德提出了蜜蜂访花偏爱性假设,此后,研究者们对各种各样的传粉昆虫也提出了访花偏爱性假设,其中包括独居蜂[30]、甲虫[31]、蝴蝶[32]和一些双翅目昆虫[33].但是,也有研究者对传粉昆虫访花偏爱性的假设理论提出了不同的见解.一种观点认为,传粉昆虫能区别和访问不同类型的花可能是由于花部特征的综合结果,包括花的颜色、形态,花瓣的大小、形状、对称性,气味等[34].Waser等(1983)假设授粉昆虫对不同花的有效利用程度和访花偏爱性程度与这些特征之间具有线性关系,而且在对蜜蜂和独居蜂的访花行为研究中发现,这两种昆虫在对花的颜色与形态学两种复合特征中所表现的偏爱性比对其中单一特征的偏爱性强.而另一种观点“特征可变性假设”认为,传粉昆虫在同一时间对复杂的花部特征信息不能进行有效的加工、处理[35],在通过多种性状来感知、确定、记忆某种花的能力上具有一定的局限性,因而在两种花存在的情况下,访花昆虫偏爱于某一单一类型的花,但这种假设只是一种理论上的推测.
本文通过长白山地区熊蜂访花偏爱性的研究,得出了熊蜂偏爱访问的蜜源植物的花部特征:具有圆锥花序、总状花序和聚伞花序的花,对于单生花,通常花形较大;访问的花冠类型以辐射对称和两侧对称为主;花多具有蜜腺,分泌花蜜;色泽艳丽,常为黄色或蓝紫色;花粉粒较大,形状多为长球形,具三孔沟,表面具网状纹饰;花的气味成分主要为酸类和酯类化合物,其中含有较高浓度的羟基丙酮、乙酸、甲酸、苯酚和9,12-十八碳二烯酸乙酯.熊蜂对于具有上述花部特征的蜜源植物表现出了偏爱性,这种偏爱性是由于蜜源植物花部特征的综合作用,还是熊蜂对某种单一类型花的特别偏爱所致,还需要进一步研究花部特征的变化与熊蜂访花行为之间的关系来加以确定.但是,可以肯定的是熊蜂的访花偏爱性表现为选择具有某些共同花部特征的蜜源植物类群.植物的花部特征与熊蜂之间存在着紧密的联系,类似的花部特征具有类似的访花昆虫类群,并且这些类群对花产生类似的选择作用.这也支持了Johnson和Steiner提出的传粉综合特征及Fenster提出的传粉者功能群的概念[36-37].
[1] BARBOSA P,LETOURNEAU D K.Natural enemies and herbivore-plant interactions:influence of plant allelochemicals and host specificity[G]//BARBOSA P,LETOURNEAU D K.Novel aspects of insect-plant interactions.New York:Wiley,1998:201-210.
[2] BELL W J,CARDÉR T.Chemical ecology of insects[M].London:Chapman and Hall,1995:433.
[3] BERNAYS E A,GRAHAM M.On the evolution of host specificity in phytophagous arthropods[J].Ecology,1998,69:886-892.
[4] BERNAYS E A.Evolution of feeding behavior in insect herbivores-success seen as different ways to eat without being eaten[J].Bioscience,1998,48:35-44.
[5] DICKE M.Chemical ecology of host-plant selection by herbivorous arthropods:a multitrophic perspective[J].Biochemical Systematics and Ecology,2000,28:601-617.
[6] 钦俊德.昆虫与植物的关系[M].北京:科学出版社,1987:12-27.
[7] HAYO H W VELTHUIS,ADRIAAN VAN DOORN.A century of advances in bumblebee domestication and the economic and environmental aspects of its commercialization for pollination[J].Apidologie,2006,37:421-451.
[8] THOMSON J D,GOODELL K.Pollen removal and deposition by honeybee and bumblebee visitors to apple and almond flowers[J].The Journal of Applied Ecology,2002,38(5):1032-1044.
[9] DAFNI A.The threat ofBombusterrestrisspread[J].Bee World,1998,79(3):113-114.
[10] SAMPSON B J,SPIERS J M,HEPP R F.Evaluating bumblebees as pollinators of Misty southern highbush blueberry growing inside plastic tunnels[J].Acta Horticulturae,2002,574:53-61.
[11] DOGTEROM M H,MATTEONI J A,PLOWRIGHT R C.Pollination of greenhouse tomatoes by the North AmericanBomnus vosnesenskii(Hymenoptera:Apidae)[J].Journal of Economic Entomology,1998,91:71-75.
[12] STOUT J C,GOULSON D.Bumble bees in Tasmania:their distribution and potential impact on Australian flora and fauna[J].Bee World,2000,81(2):80-86.
[13] BUTTERMORE R E.Observation of successfulBombusterrestris(L.)(Hymenoptera:Apidae)colonies in southern Tasmania[J].Australian Journal of Entomology,1997,36:251-254.
[14] Donovan B J.Interacions between native and introduced bees in New Zealand[J].New Zealand Journal of Ecology,1980(3):104-116.
[15] ABAK K,OZDOGAN A O,DASGAN H Y,et al.Effectiveness of bumble bees as pollinators for eggplants grown in unheated greenhouses[J].Acta Horticulturae,2000,514:197-203.
[16] 安建东,李磊,孙永深,等.熊蜂为温室西红柿授粉的效果研究[J].蜜蜂杂志,2001(9):13-15.
[17] 安建东,邢艳红,彭文君,等.日光温室桃园释放熊蜂授粉试验[J].中国果树,2003(5):13-14.
[18] 安建东,童越敏,国占宝,等.熊蜂为温室茄子授粉试验[J].中国养蜂,2004,55(3):7-8.
[19] 国占宝,李乃光,孙永深.熊蜂为温室黄瓜授粉的效果研究[J].蜜蜂杂志,2002(6):3-4.
[20] 孙永深,安建东,童越敏,等.熊蜂为温室冬瓜授粉的效果研究[J].蜜蜂杂志,2003(8):3-5.
[21] 李乃光,彭文君,安建东.熊蜂为温室蔬菜授粉技术应用[J].蜜蜂杂志,2002(11):30-31.
[22] 龚禹峰,邢艳红,马书英,等.熊蜂为温室大棚桃授粉技术应用[J].中国养蜂,2001,52(5):18-21.
[23] HEINRICH B,RAVEN P H.Energetics and pollination ecology[J].Science,1972,176:597-602.
[24] PELLMYR O.Pollination by animals[G]// HERRERA C M,PELLMYR O.Plant-animal interactions:an evolutionary approach.Oxford:Blackwell Science Ltd,2002:157-184.
[25] BARRETT S C H,HARDER L D.Ecology and evolution of plant mating[J].Trends in Ecology and Evolution,1996,11:73-79.
[26] 尚利娜.长白山区访花昆虫与蜜源植物的协同适应[D].长春:东北师范大学,2010.
[27] 官昭瑛.长白山地区访花昆虫访花机制研究[D].长春:东北师范大学,2006.
[28] ERDTMAN G.Handbook palynology[M].Beijing:Science Press,1969:34-56.
[29] ROBERT J G,TERENCE M L.Flower constancy in bumblebees:a test of the trait variability hypothesis[J].Animal Behaviour,2003,69,939-949.
[30] GROSS C L.Floral traits and pollinator constancy:foraging by native bees among three sympatric legumes[J].Australian Journal of Ecology,1992,17:67-74.
[31] DE LOS MOZOS PASCUAL M,DOMINGO L.Flower constancy inHeliotaurusruficollis(Fabricius,1781),Coleoptera,Alleculidae[J].Elytron,1991(5):9-12.
[32] LEWIS A C.Memory constraints and flower choice inPierisrapae[J].Science,1986,232:863-865.
[33] GOULSON D,WRIGHT N P.Flower constancy in the hoverfliesEpisyrphusbalteatus(Degeer)andSyrphusribesii(L.)(Syrphidae)[J].Behavioral Ecology,1998(9):213-219.
[34] WASER N M.The adaptive nature of flower traits:ideas and evidence[G]//REAL L A.Pollination biology.Orlando,Florida:Academic Press,1983:241-285.
[35] GEGEAR R J,LAVERTY T M.The effect of variation among floral traits on the flower constancy of pollinators[G]//CHITTKA L,THOMSON J D.Cognitive ecology of pollination:animal behaviour and floral evolution.Cambridge:Cambridge University Press,2001:1-20.
[36] JOHNSON S D.STEINER K E.Generalization versus specialization in plant pollination systems[J].Trends in Ecology and Evolution,2000,15:140-143.
[37] FENSTER C B,ARMBRUSTER W S,WILSON P,et al.Pollination syndromes and floral specialization[J].Annual Reviews of Ecology,Evolution and Systematics,2004,35:375-403.
The study of preference of flower-visiting bumblebees in Changbai Mountain Region
REN Bing-zhong,SHANG Li-na,CHEN Xin,HAN Ye,XU Yan
(School of Life Sciences,Northeast Normal University,Changchun 130024,China)
This paper studied the flower shape,color,scent and pollen morphology of main nectar plants visited by seven bumblebees in Changbai Mountain Region,and drew the conclusion of the floral characteristics preferred by bumblebees.Bumblebees preference for a visit of flowers with panicles,raceme and cyme,and for a single flower,usually with a larger flower-shaped;corolla types are mainly radial symmetry and bilateral symmetry;mostly with nectary to secret nectar;colorful,and often yellow or blue purple;pollen large,mostly prolate shape,with three colporates,reticulate surface;flower scent was mainly acids and esters,which contained a higher concentration of 1-hydroxy-2-propanone,acetic acid,formic acid,phenol and 9,12-octadecadienoic acid,ethyl ester.This study will provide the basic material for bumblebee pollination and the application of new agents to lure pollinating insects.
Changbai Mountain Region;Bombus;preference of flower-visiting
Q 969.557.7
180·5430
A
1000-1832(2012)01-0111-07
2011-07-23
吉林省科技发展计划项目(20040547-1).
任炳忠(1963—),男,博士,教授,博士研究生导师,主要从事昆虫学研究.
方 林)
猜你喜欢
中国科学探险(2022年8期)2022-12-31
疯狂英语·新悦读(2022年12期)2022-03-08
今日农业(2021年6期)2021-06-09
新疆大学学报(自然科学版)(中英文)(2020年2期)2020-07-25
青少年科技博览(中学版)(2020年1期)2020-04-21
奥秘(创新大赛)(2019年4期)2019-04-15
儿童故事画报·发现号趣味百科(2017年10期)2018-03-13
儿童故事画报·发现号趣味百科(2017年9期)2018-03-13
儿童故事画报·发现号趣味百科(2016年4期)2016-06-24
奥秘(2015年3期)2015-09-10
