
波斯菊核型分析
2012-12-24 00:53:38龙双红
草业科学 2012年11期
张 凡,张 芹,龙双红
(1.河北农业大学园林与旅游学院,河北 保定071001;2.河北省塞罕坝机械林场总场,河北 围场068466)
波斯菊(Cosmosbipinnatus)原产于墨西哥,为菊科秋英属一年生草本花卉,其叶形雅致,花色丰富,花期长,抗逆性强,栽培管理容易,是优良的园林绿化材料,常用作地被植物,也可用于花境、花丛或建筑墙垣的基础栽植。目前有关波斯菊的研究主要集中在播种、育苗、栽培、应用等方面[1-3],而对于染色体核型的研究还比较少。Thomas[4]曾报道墨西哥部分波斯菊的染色体数目,杨德奎和周俊英[5]报道过山东部分地区波斯菊的核型,但仍缺乏不同地区不同环境条件下波斯菊染色体及核型更广泛的研究。本研究对生长在海拔较高的塞罕坝地区的波斯菊染色体观察、计数及核型分析,并与前人的研究进行比较,旨在为波斯菊的种质鉴定、起源演化及品种改良补充更有力的证据。
1 材料与方法
1.1 材料 供试波斯菊种子于2010年9月采自河北省塞罕坝机械化林场,置于4℃冰箱,保存备用。
1.2 方法 取饱满的种子,用蒸馏水浸泡12h后,在25℃恒温箱中培养。当幼根长约1cm时,选取根毛粗壮、根尖白嫩的幼根用0.002mol·L-1的8-羟基喹啉在20℃恒温箱中预处理2h,然后转入卡诺固定液20℃固定24h。取出根尖,用蒸馏水冲洗3~5遍后,放入1mol·L-1的HCl在60℃水浴中解离6min。取解离好的幼根分生组织放在载玻片中央,加1滴铁矾-苏木精染色剂,盖盖玻片,在酒精灯上加热数秒,再轻压使材料尽量分散。然后在Olympus BH22光学显微镜下观察,选择染色清晰、染色体分散而不重叠的分裂中期细胞进行染色体数目统计并拍照[6]。
染色体计数和核型分析依据李懋学和陈瑞阳[7]的分析标准进行,统计分散较好的30个以上分裂中期细胞,以85%以上的细胞具有相同的染色体数作为该种的染色体数,并选取分散良好的5个中期分裂相进行分析测量,取平均值计算核型数据。
核型类型分析参照Stebbins[8]的分类标准,染色体类型分析按照Levan等[9]的分类系统。核型不对称系数按Arano[10]的方法计算,比值越大越不对称。
2 结果与分析
2.1 波斯菊的染色体数目 选择分散良好的30个中期分裂相的细胞计数,其中28个细胞染色体数目为24条(图1),占计数细胞的93.3%。根据染色体的长度、臂比、着丝粒的位置、随体的有无和位置等进行同源染色体配对,每个细胞的24条染色体可配成12对,表明均为二倍体,染色体基数为n=12。
2.2 波斯菊染色体核型分析 预处理及固定条件不同会直接影响染色体的短缩程度,所以即使是同一植物,不同研究所得到的绝对长度数值也会有明显差异,而相对长度则是一组可比性更高的、稳定的数值。因此,本研究通过精确测量5个分散良好的中期分裂相的细胞染色体的长臂、短臂的相对长度,并通过计算得到波斯菊染色体的核型参数。在波斯菊12对同源染色体中,第6、8和11号染色体的臂比在1.71~3.00,为近中着丝粒染色体(sm型),占染色体总数的25%;其余染色体的臂比在1.01~1.70,为中间着丝粒染色体(m型),占染色体总数的75%(表1)。本研究发现2对具有随体的染色体,分别为6和12号,随体在染色体短臂的近端部(图2)。因此,可以确定波斯菊的染色体核型公式为2n=2x=24=18m(1sat)+6sm(1sat)。

图1 波斯菊的染色体及核型Fig.1 Chromosome and karyotype of Cosmos bipinnatus
波斯菊染色体由14条中长染色体(M2)和8条中短染色体(M1)及2条短染色体(S)组成,染色体相对长度平均值在6.20%~9.06%(表1)。根据Stebbins[8]的“不对称核型分析”分类标准,波斯菊最长与最短的染色体长度比值为1.46,臂比大于2∶1的染色体为2对,占染色体总数的16.7%,应属于“2A”型;核型不对称系数为43.25%,对称程度较高,为较对称类型。

图2 波斯菊的染色体核型模型图Fig.2 Karyotype diagram of Cosmos bipinnatus
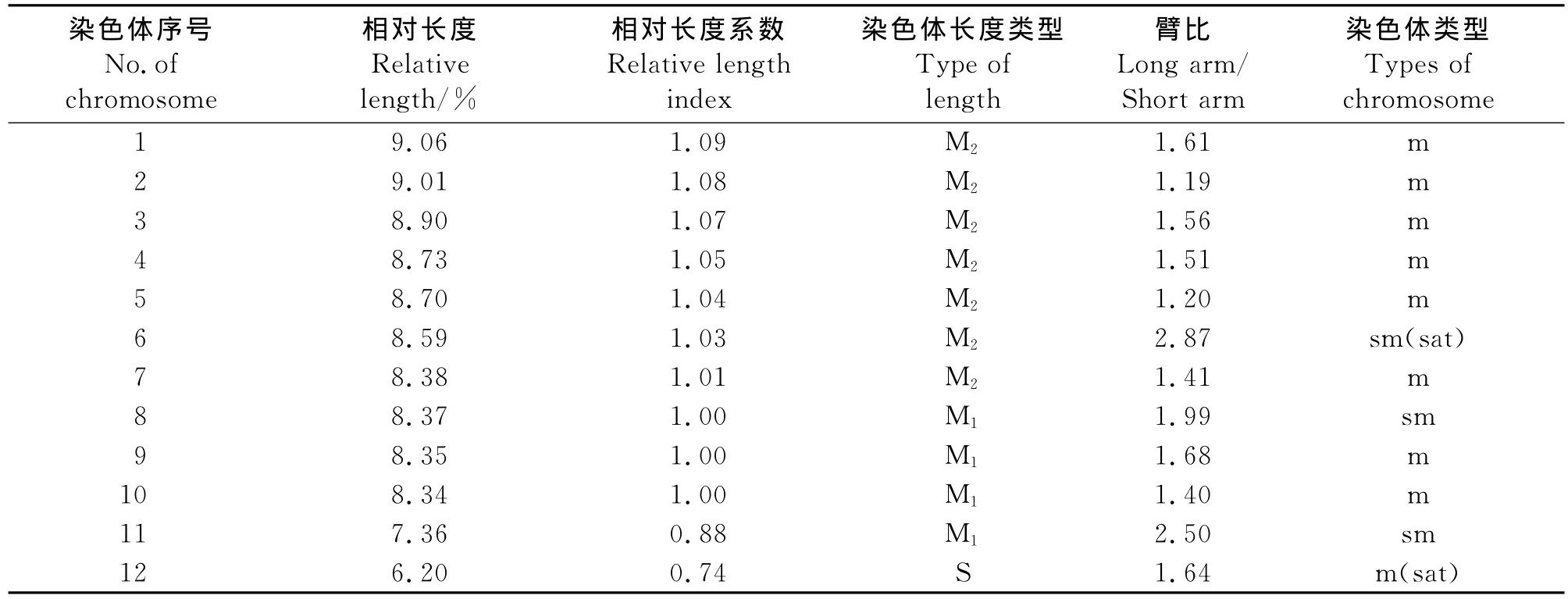
表1 波斯菊的核型参数Table 1 Karyotype index of Cosmos bipinnatus
3 讨论
菊科秋英属植物全世界约25种,我国常见栽培的有波斯菊和硫华菊(C.sulphureus)两种[11]。国外有关该属植物染色体数目报道的有10种左右,染色体基数分为24和12两类[4]。本研究中波斯菊的染色体基数为12,与国外的报道一致,也与杨德奎和周俊英[5]研究报道的染色体数目相同,虽然以上研究取材的地点差异很大,但其染色体数目受环境影响较小,具有很高的稳定性和再现性,因此可以确定波斯菊为二倍体,有24条染色体。
本研究与杨德奎和周俊英[5]的报道一致,认为波斯菊的核型类型为“2A”型,属于较对称类型。而与其同属的硫华菊的染色体核型公式为2n=24=2sm+22st,核型类型为“4B”型,是很不对称的核型[12]。根据高等植物染色体的进化理论[8],可以确定波斯菊比同属的硫华菊更原始。
山东地区的波斯菊的核型公式为2n=24=16m+2m(sat)+6sm,其染色体只有1对带随体[5],而本研究取材于塞罕坝的波斯菊的染色体中有2对带随体,分别是中部着丝粒染色体(m)和近中部着丝粒染色体(sm),并且染色体的长度与前人报道也有一定差异。这可能与生境不同有关,在其他种的染色体研究中也有不同居群核型差异的报道[13-15],要证实这一结论还需要扩大取样地点进一步研究,甚至有必要结合分子生物学的手段进行深入研究。另外,染色体长度在预处理过程中受药剂种类、浓度及处理时间的影响较大,试验中所采取的处理方法、观察手段的差别,也可能是导致试验结果不同的原因之一,在今后的研究中应进一步优化方法,提高核型分析的精确度。
致谢:对河北农业大学园艺学院张成合老师在制片过程中给予的指导和帮助表示感谢!
[1]杜寿辉,徐涛,何素瑞,等.昆明地区波斯菊植物结实率及种子萌发特性研究[J].安徽农业科学,2010,38(28):15493-15494.
[2]潘永飞,戴忠良,潘叶飞,等.播种期对波斯菊结实和种子发芽率的影响[J].金陵科技学院学报,2009,25(2):64-67.
[3]张娟,杨玉想.大花波斯菊在北方地区的栽培与应用[J].现代园艺,2010(9):20.
[4]Thomas E M.Systematic studies in the Coreopsidinae:Cytotaxonomy of Mexican and Guatemalan Cosmos[J].American Journal of Botany,1968,55(3):345-353.
[5]杨德奎,周俊英.山东10种植物的核型分析[J].广西植物,1999,19(4):349-354.
[6]李懋学,张赞平.作物染色体及其研究技术[M].北京:中国农业出版社,1996:5-17.
[7]李懋学,陈瑞阳.关于植物核型分析的标准化问题[J].武汉植物学研究,1985,3(4):297-302.
[8]Stebbins G L.Chromosomal Evolution in Higher Plants[M].London:Edward Arnold,1971:85-104.
[9]Levan A,Fredga K,Sandberg A A.Nomenclatrue for centromeric position of chromosomes[J].Hereditas,1964,52:201-220.
[10]Arano H.Cytological studies in subfamily carduoideae(Compositae)of Japan,IX[J].Botannical Magazine(Tokyo),1963,76:32-40.
[11]贺士元.河北植物志(第三卷)[M].石家庄:河北科学技术出版社,1991:75-76.
[12]汪小兰,李懋学.10种菊科花卉的染色体观察[J].武汉植物学研究,1987,5(2):111-117.
[13]时丽冉,高汝勇,李会芬,等.紫茉莉染色体数目及核型分析[J].草业科学,2010,27(1):53-54.
[14]张晓燕,毛培春,孟林,等.三份偃麦草种质的染色体核型分析[J].草业学报,2011,20(4):194-200.
[15]周自玮,奎嘉祥,钟声,等.云南野生鸭茅的核型研究[J].草业科学,2000,17(6):48-51.
猜你喜欢
生物技术进展(2022年1期)2022-02-11 03:45:46
河北农业大学学报(社会科学版)(2021年2期)2021-06-24 05:59:08
云南化工(2020年11期)2021-01-14 00:50:46
诗选刊(2020年4期)2020-04-09 04:49:43
科学之谜(2018年9期)2018-12-17 09:05:08
黄河之声(2018年10期)2018-07-15 07:34:46
西藏农业科技(2018年4期)2018-04-25 06:39:38
文学港(2018年1期)2018-01-25 12:48:15
绿色中国(2017年19期)2017-05-25 13:25:15
生物学教学(2016年12期)2016-04-10 23:04:28
