
青藏高原东缘15种藜科和蓼科植物种子萌发特性
2012-12-24 00:53吕俊平张春辉张莹莹杜国祯王晨阳
草业科学 2012年2期
吕俊平,张春辉,张 蕾,张莹莹,杜国祯,王晨阳
(1.兰州大学干旱与草地生态教育部重点实验室,甘肃 兰州730000;2.兰州大学草地农业科技学院,甘肃 兰州730020)
种子萌发行为作为植物生活史对策的主要内容,已受到生态学家极大关注[1-7]。种子的萌发特征影响着幼苗存活能力[8]、个体适合度[9-10]和植物生活史的表达[11]。因此,对种子萌发行为的研究已成为植物生态学研究的重要课题。
光照作为一种重要的环境因子,对种子萌发具有重要影响。为防止种子在不利于幼苗建植的地点与时间萌发,种子萌发对光照的响应至关重要[12]。光对种子萌发的影响主要是作为一种刺激信号,打破种子的休眠,而不是作为一种能量物质直接参与。Vazquez-Yanes和Orosco-Segovia[13]依据种子萌发对光敏感程度的不同将萌发分为以下3类:1)种子仅能在光照条件下萌发;2)种子仅能在黑暗条件下萌发,光抑制萌发;3)种子在光照和黑暗条件下都能萌发。
种子萌发是一种极其复杂的现象,它不仅与环境因子相关而且与种子特有的属性也有着密切的关系[14]。种子大小与种子萌发率呈负相关[15],与幼苗的存活率呈正相关[16]。因此,种子大小能够影响种子的萌发能力,并且直接影响幼苗的建植,最终影响整个群落的结构[17]。此外,不同植物区系中的研究结果均表明种子大小与生活型相关[18]。Moles等[19]研究了12 987种植物的种子大小数据,表明植物的生活型是影响种子大小的最重要的一个特征。相对于多年生植物,一年生植物具有较小的种子[20-21]。一年生植物种子质量小于二年生和多年生植物的种子质量[17]。因此,可以推测生活型也可能影响种子的萌发能力。
综上所述,通过研究不同光照条件下的种子萌发行为,拟解决以下两个问题:1)种子大小是否对种子萌发对光照的响应产生影响?2)生活型是否也对种子萌发对光照的响应产生影响?关于萌发对光照响应的研究大多集中于热带、新热带木本以及欧洲温带森林草本中[22-25]。而在高寒地区,尤其是青藏高原地区,此类研究还处于空白。由于藜科与蓼科植物在甘南分布较为广泛[26-27]而且以一年生和多年生两种生活型为主。因此,本研究选取15种藜科和蓼科植物,旨在研究该地区蓼科和藜科植物种子萌发对光照响应的规律,以及种子大小和生活型是如何对种子萌发对光照的响应产生影响的。
1 材料与方法
1.1 研究区自然概况 本研究所用15种藜科和蓼科植物种子采自甘肃省甘南藏族自治州境内,研究区位于青藏高原东部地区(101°~103°E,34°~35°70′N),海拔2 900~4 000m;年均降水量450~780mm,降水主要分布7―9月;年均气温1.8℃,1月均温-10.7℃,7月均温11.7℃,生长季最高温28.9℃;年平均霜期大于270d。萌发试验在兰州大学高寒草甸与湿地生态系统定位研究站(合作市,34°55′N,102°53′E)进行;该地海拔2 942m;年平均气温2.0℃;年均降水量550mm。植被类型为亚高寒草甸,常见物种有披碱草(Elymussp.)、鹅观草(Roegneriasp.)、羊茅(Festucasp.)和银莲花(Anemonesp.)等。
1.2 试验种子处理及称量 将采集来的种子在实验室内风干并去除杂质,然后装在信封中室温下储藏(温度约为15℃)。将每种植物的种子混合在一起,从中随机选取3份饱满种子(每份100粒),并用万分之一的电子天平称量百粒重。种子质量定义为胚、胚乳加上种皮的质量。
1.3 物种描述 本试验所选植物为5种藜科植物和10种蓼科植物,其中一年生为9种,多年生为6种。以种子质量逐渐增加的方向对物种进行了编号(表1)。

表1 物种名录Table 1 List of plant species in present study
1.4 萌发试验 2009年6月,在试验站内用黑色尼龙网眼布(遮阳网)搭建遮阳棚,并设置了2个光照梯度处理,其光照强度分别为全自然光照(100%Natural Light,NS)和自然光强的3%(2层遮阳网搭建)。不同光照强度下,每个物种设置3个重复,每个重复随机选取100粒饱满种子。进行萌发试验时,在直径为17cm的花盆内装填10cm深的土,并在土表铺一张白色纯棉布(防止花盆装填土壤中的种子库种子发芽影响试验结果),然后将种子均匀放置在棉布上(放置有种子的棉布上不覆土,以使种子与外界环境条件充分接触)。放入种子后,定期浇水,使棉布保持湿润。每天统计萌发个数,胚根长度等于种子长度时记为萌发,并将已经萌发的种子移出花盆。萌发试验持续114d。
1.5 数据分析
1.5.1 萌发指标 萌发率(Germination Percentage,GP)=种子发芽总数/供试种子数(100粒)×100%,表示萌发周期结束后,萌发种子的数量占总供试种子的比例,反映种子的萌发数量。
相对光萌发率(Relative Light Germination,RLG)=Gl/(Gd+Gl)[23],计算种子萌发对光的依赖程度,式中,Gl为全光照下的萌发率,Gd为黑暗条件下的萌发率。RLG在0(越接近1,表示种子倾向于在低光照或者黑暗条件下达到较大的萌发率)到1(越接近1,表示种子倾向于在高光照条件下达到较大的萌发率)之间变化。
本研究设计的光照梯度为自然光强的3%和100%。因此,RLG表示在3%~100%的自然光强下种子萌发对光的依赖程度。
1.5.2 数据处理 以配对T检验分析光照强度对种子萌发能力影响,以Pearson相关分析评价物种相对光萌发率与种子大小的关系。为了确保方差齐性,在统计分析之前,对种子大小进行了对数转换。所有分析均在 SPSS 16.0(SPSS Inc.,Chicago,USA)中完成。
2 结果与分析
2.1 藜科和蓼科植物种子大小的分布 本研究所选取的15种藜科和蓼科植物,其百粒重为16.53~2 174.17mg,平均为263.40mg,种子大小变异跨越了2个数量级(图1),最小的是青藏蓼,为16.53mg,最大的是苦荞麦,为2 174.17mg。13个物种的百粒重小于263.40mg,占总种数的86.67%。将种子百粒重进行对数转换后,种子大小频数呈正态分布。
2.2 不同光照水平下藜科和蓼科植物种子的萌发率 在不同光照水平下,此15种植物种子的萌发率各不相同,但大体上可以分为以下两类:1)在2个光照水平下,土荆芥、猪毛菜、藜、披针叶酸模、轴藜、扁蓄、尼泊尔酸模的萌发率无显著差异(P>0.05);2)青藏蓼、灰绿藜、冰川蓼、皱叶酸模、圆穗蓼、水生酸模、巴天酸模、苦荞麦的萌发率差异显著(P<0.05)。其中,青藏蓼、冰川蓼、圆穗蓼、苦荞麦在3%自然光照下的萌发率显著高于100%自然光照下的萌发率,光照抑制了萌发;而灰绿藜、皱叶酸模、水生酸模、巴天酸模在100%自然光照下的萌发率显著高于3%自然光照下的萌发率,光照促进了萌发(图2)。

图1 种子大小分布Fig.1 Distribution of seeds size
2.3 藜科和蓼科植物相对光萌发率与种子大小之间的关系 本研究表明,整体上,RLG与种子大小间呈显著负相关关系(r=-0.303,P=0.043),即随着种子大小的增加,种子萌发对光的依赖性降低(图3)。生活型方面,一年生植物,RLG与种子大小间呈显著负相关(r=-0.679,P<0.001),即随着种子大小的增加,种子萌发对光照的依赖性显著降低;而多年生植物,RLG与种子大小间无相关关系(P>0.05),即多年生植物种子萌发不依赖于光照(图3)。

图2 不同光照水平下15种植物种子萌发率差异Fig.2 Difference of seed germination rates of 15plant species under different light condition
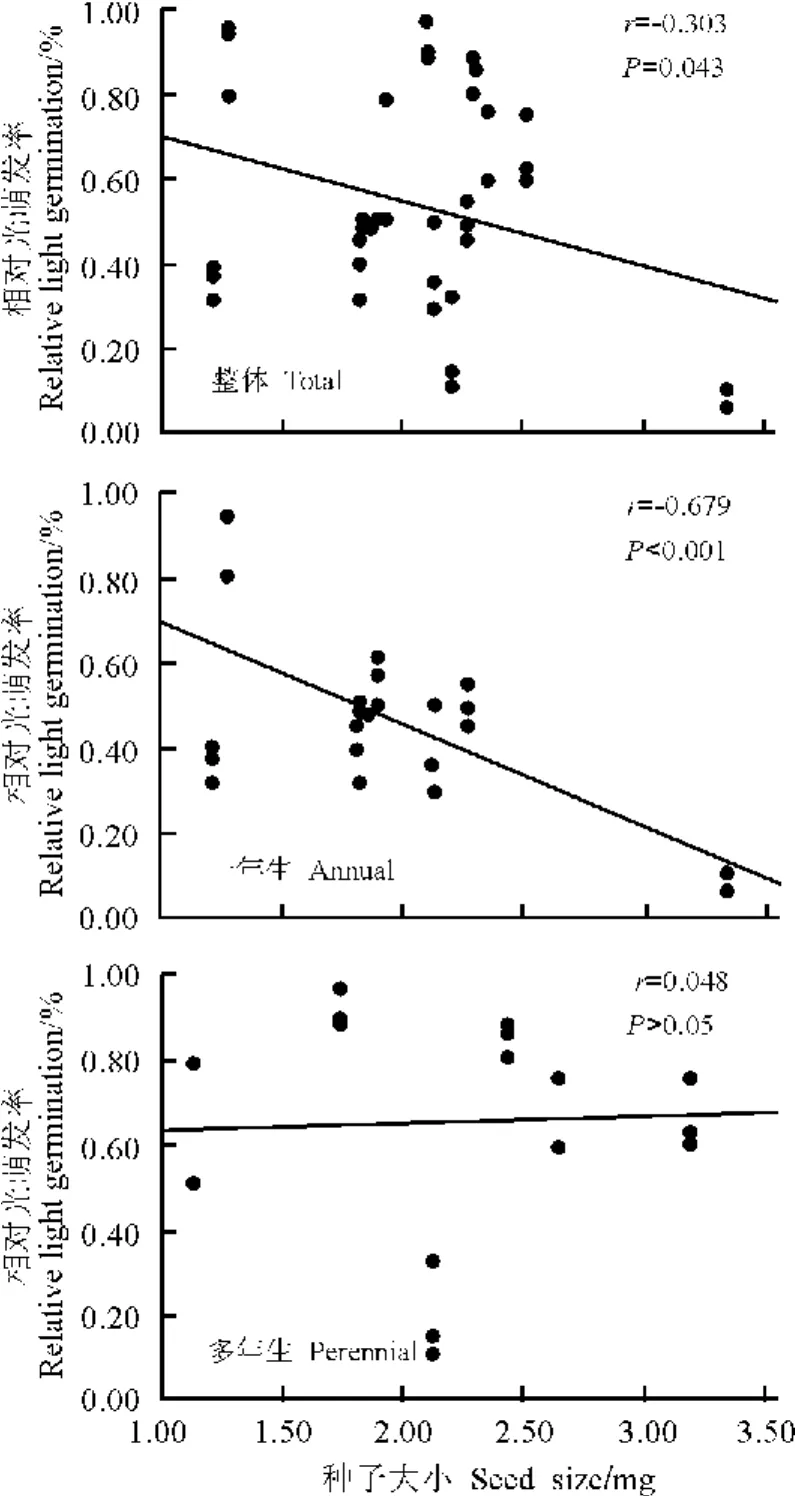
图3 藜科和蓼科植物相对光萌发率与种子大小的关系Fig.3 Relationship between seed size and relative light germination of Chenopodiaceae and Polygonaceae species
3 讨论
3.1 藜科和蓼科植物相对光萌发率与种子大小之间的关系 Vazquez-Yanes和 Smith[28]认为,临窗环境中光照条件的差异是影响种子萌发最为直接的因子。本研究所选取的15种植物,就单个物种而言,萌发对光照的响应各不相同,这可能与它们所生长的微生境下的光照条件密切相关。而整体上,小种子比大种子更易对光照产生响应,即随着种子大小的增加,种子萌发对光的依赖性逐渐降低。Milberg等[23]对54种欧洲草本种子萌发对光响应的研究认为,随着种子大小的增加,种子萌发对光的依赖性逐渐降低。Pearson等[24]认为,种子萌发对光的响应与种子大小密切相关。此结论可能是种子萌发对光响应的一般模式,可以从以下两个方面作出解释。1)种子大小作为植物生活史的一个关键特征,它影响着幼苗的建植、存活及种子扩散等更新对策。种子大小反映了幼苗初始所能获取的资源量的多少[29],而且幼苗的存活依赖于种子所供给的养分[30-31]。小种子由于储藏的营养物质较少,仅仅能够在较浅的土壤表层出土[32];大种子具有较多的资源,能够从较深的土层中成功出苗[21]。而光仅能够穿透几毫米的土层[33]。因此,需光萌发作为深度感应机制能够确保小种子仅能从较浅的土层中萌发。2)通常,种子大小反映了幼苗的初始大小。大种子的幼苗通常比小种子的幼苗大[30]。因此,大种子有利于幼苗较早地发展成为完善的资源捕获系统(根系组织和光合组织)[34-35]。而小种子幼苗,尤其在低光照条件下,其初始大小较小并且具有较小的胚根生长速率,这种状态将持续较长的一段时间。因此,就幼苗个体而言,小种子幼苗较脆弱,并且将面临包括践踏、干旱、植食和埋藏等风险[29,36]。青藏高原东缘海拔较高,寒冷湿润,年平均温度较低,生长季短,霜害等自然灾害频繁,各种环境因子的变化具有随机性和无法预测性。因此,为了降低死亡风险,提高幼苗存活率,一旦遇到适宜的光照条件,小种子便快速萌发并且通过高光照条件下的幼苗快速生长来缩短“小幼苗”阶段。小种子的这种对光照的敏感性是其对环境适应的必然结果。
3.2 生活型与种子萌发对光响应的关系 不同植物区系中的研究结果均表明种子大小与生活型相关[18,20]。Baker[20]、Leishman等[21]发现,一年生植物具有较小的种子。Silvertown[17]也发现一年生植物种子质量小于二年生和多年生植物的种子质量。因此,本研究将植物分为一年生与多年生两种生活型。一年生植物种子萌发对光照敏感,而多年生植物种子萌发不依赖于光照。多年生植物具有较大的种子,因而增加了扩散后被捕食的风险[37-38]。在青藏高原东缘,草地生态系统承载着大量牛、羊等牲畜,且生存着许多以植物根茎及其种子为食物的啮齿类动物,因此,较大的种子类群极易成为被捕食的目标。由于大种子能够从较深的土层中出苗,可以通过较深的埋藏深度来降低被捕食风险[24],而光只能够穿透几毫米的土壤层,不能到达大种子所处的土壤深度。所以,大种子植物的萌发不依赖于光照。以往的研究结果[24]也表明,大种子植物的萌发对温度波动较为敏感。
3.3 种子大小相关的萌发对光响应的另一种解释机制 依据种子大小数目权衡模型[39],植物的种子大小与其所能产生的种子数目呈负相关,与幼苗的存活率呈正相关。小种子植物繁殖产生大量的小种子,通过扩散等方式拓殖到许多领域,其中包括大种子植物不能到达的领域。而大种子植物繁殖产生少量竞争力强的大种子。因此,小种子为了增加适应力,必须尽可能多地占领适宜萌发的斑块,并在斑块中迅速大量萌发来增加幼苗成活率,进而增加整体适应力。种子萌发对光照的正响应是种子寻找斑块的一种机制,因为斑块中的光照强度比植被下面要强的多。小种子萌发能对光做出正响应,保证小种子尽可能的在竞争强度较低的斑块中萌发,促进了小种子植物的幼苗成活率,有利于小种子植物的建植。而大种子能够在其所在的领域成功建植,产生较大的幼苗,有较强的竞争能力,对不利环境的耐受性也较强。因此,大种子萌发对光照强度的变化并不十分敏感。大种子物种为遮阴耐受型物种,其较为丰富的储藏物质能够为其根系组织和光合组织的发展提供充足的物质基础[40]。相对于小种子植物,大种子植物的幼苗能够汲取更多的养分和捕获更多的光照。因此,大种子植物的种子能够在较低的光照强度下萌发并且对光照强度的变化并不十分敏感。
4 结论
整体上,随着种子大小的增加,种子萌发对光照的依赖性逐渐降低。一年生植物种子萌发对光照较为敏感;而多年生植物种子萌发不依赖于光照。种子大小和生活型共同影响着种子萌发对光照的响应。
[1]刘志民,李雪华,李荣平,等.科尔沁沙地31种一年生植物萌发特性比较研究[J].生态学报,2004,24(3):648-653.
[2]罗亚勇,赵学勇,黄迎新,等.三种一年生藜科沙生植物出苗对沙埋深度和水分条件的响应[J].草业科学,2009,18(2):122-129.
[3]李巧峡,李凯,丁文龙,等.不同处理对北方嵩草种子萌发的影响[J].草业科学,2009,26(8):112-117.
[4]黄文达,王彦荣,胡小文.三种荒漠植物种子萌发的水热响应[J].草业学报,2009,18(3):171-177.
[5]王梅英,刘文,刘坤,等.青藏高原东缘10种禾本科植物种子萌发的基温和积温[J].草业科学,2011,28(6):983-987.
[6]王进,张勇,颜霞,等.光照、温度、土壤水分和播种深度对披针叶黄华种子萌发及幼苗生长的影响[J].草业科学,2011,28(9):1640-1644.
[7]郭淑青,齐威,王玉林,等.青藏高原东缘海拔对植物种子大小的影响[J].草业学报,2011,19(1):50-58.
[8]Gross K L,Smith A D.Seed mass and emergence time effects on performance ofPanicumdichotomiflorumMichx across environments[J].Oecologia,1991,87:270-278.
[9]Donohue K,Dom L,Griffith C,etal.The evolutionary ecology of seed germination ofArabidopsisthaliana:variable natural selection on germination timing[J].E-volution,2005,59:758-770.
[10]Miller T.Effects of emergence time on survival and growth in an early old-field plant community[J].Oecologia,1987,72:272-278.
[11]Venable D L.The evolutionary ecology of seed heteromorphism[J].The American Naturalist,1985,126:577-595.
[12]Fenner M,Thompson K.The Ecology of Seeds[M].Cambridge:Cambridge University Press,2005:116-117.
[13]Vazquez-Yanes C,Orosco-Segovia A.Patterns of seed longevity and germination in the tropical rainforest[J].Annual Review of Ecology and Systematics,1993,24:69-87.
[14]Baskin C C,Baskin J M.Seeds[M].San Diego:Academic Press,1998:13-16.
[15]Bu H Y,Chen X L,Xu X L,etal.Seed mass and germination in an alpine meadow on the eastern Tsinghai-Tibet plateau[J].Plant Ecology,2008,191:127-149.
[16]Peco B,Traba J,Levassor C,etal.Seed size,shape and persistence in dry Mediterranean grass and scrublands[J].Seed Science Research,2003,13:87-95.
[17]Silvertown J W.Seed size,life span and germination date as co-adapted features of plant life history[J].The American Naturalist,1981,118:860-864.
[18]Thompson K,Rabinowitz D.Do big plants have big seeds[J].The American Naturalist,1989,133(5):723-728.
[19]Moles A T,Ackerly D D,Webb C O,etal.A Brief history of seed size[J].Science,2005,307:576-580.
[20]Baker H G.Seed weight in relation to environmental conditions in California[J].Ecology,1972,53(6):997-1010.
[21]Leishman M R,Westoby M,Jurado E.Correlated of seed size variation——A comparison among five temperate floras[J].Journal of Ecology,1995,83:517-529.
[22]Metcalfe D J,Grubb P J,Turner I M.The ecology of very small-seeded shade-tolerant trees and shrubs in lowland rain forest in Singapore[J].Plant Ecology,1998,134:131-149.
[23]Milberg P,Andersson L,Thompson K.Large seeded species are less dependent on light for germination than small-seeded ones[J].Seed Science Research,2000,10:99-104.
[24]Pearson T R H,Burslem D F R P,Mullins C E,etal.Germination ecology of neotropical pioneers:Interacting effects of environmental conditions and seed size[J].Ecology,2002,83:2798-2807.
[25]Pearson T R H,Burslem D F R P,Mullins C E,etal.Functional significance of photoblastic germination in neotropical pioneer trees:a seed’s eye view[J].Functional Ecology,2003,17:394-402.
[26]常朝晖.甘肃蓼科饲用植物资源研究[J].甘肃科技,2007,23(4):222-223.
[27]王一峰,王俊龙.甘肃藜科植物研究[J].植物研究,2006,26(6):742-749.
[28]Vazquez-Yanes C,Smith H.Phytochrome control of seed germination in the tropical rain forest pioneer treesCecropiaobtusifoliaandPiperauritumand its ecological significance[J].New Phytologist,1982,92:477-485.
[29]Leishman M R,Wright I J,Moles A T,etal.The E-cology of Regeneration in Plant Communities[M].2nd edition.Wallingford:CABI Publishing,2000:237-256.
[30]Harper J L.Population Biology of Plants[M].London:Academic Press,1977:26-31.
[31]Fenner M.Seed Ecology[M].New York:Chapman and Hall,1985:49-53.
[32]Bond W J,Honig M,Maze K E.Seed size and seedling emergence:An allometric relationship and some ecological implications[J].Oecologia,1999,120:132-136.
[33]Tester M,Morris C.The penetration of light through soil[J].Plant,Cell and Environment,1987,10:281-286.
[34]Armstrong D P,Westoby M.Seedlings from large seeds tolerate defoliation better:A test using phylogenetically independent contrasts[J].Ecology,1993,74:1092-1100.
[35]Foster S A.On the adaptive value of large seeds for tropical moist forest trees:A review and synthesis[J].The Botanical Review,1986,52:260-299.
[36]Daws M I,Pearson,T R H,Burslem D F R P,etal.Effects of topographic position,leaf litter and seed size on seedling demography in a semi-deciduous tropical forest in Panama[J].Plant Ecology,2005,179:93-105.
[37]Blate G M,Peart D R,Leighton M.Post-dispersal predation on isolated seeds:A comparative study of 40tree species in a southeast Asian rainforest[J].Oikos,1998,82:522-538.
[38]Kollmann J,Coomes D A,White S M.Consistencies in post-dispersal seed predation of temperate fleshyfruited species among seasons,years and sites[J].Functional Ecology,1998,12:683-690.
[39]Smith C C,Fretwell S D.The optimal balance between size and number of offspring[J].The American Naturalist,1974,108:499-506.
[40]Nina H.Seed size and shade-tolerance:A comparative analysis of North American temperate trees[J].Oecologia,1998,114:432-440.
猜你喜欢
制造技术与机床(2019年6期)2019-06-25
小学生学习指导(低年级)(2018年9期)2018-09-26
读者·校园版(2018年1期)2017-12-16
成才之路(2017年21期)2017-07-27
安徽农学通报(2017年9期)2017-05-19
现代农业科技(2016年22期)2017-03-24
现代制造技术与装备(2015年4期)2015-12-23
浙江大学学报(农业与生命科学版)(2015年4期)2015-12-15
原子与分子物理学报(2015年1期)2015-11-24
少年科学(2015年2期)2015-08-12
