
植物硒转运蛋白的研究进展
2012-12-23 04:05马建辉孙梅好
山西农业科学 2012年8期
马建辉,孙梅好
(浙江师范大学化学与生命科学学院,浙江金华321004)
硒(selenium,Se)是哺乳动物、细菌等许多生物的必需微量元素[1-3],适量补充硒有利于植物的生长发育,可提高农作物的产量及品质[4-8]。硒在土壤中的平均质量分数大约为0.4 mg/kg,某些地区的硒含量超过10 mg/kg[9],可能会导致动物以及人类硒中毒[10-12]。人体内已发现25 种硒蛋白(selenoprotein),例如谷胱甘肽过氧化物酶(glutathione peroxidase,GPx)、硫氧还蛋白还原酶(thioredoxin reductase,TrxR)、甲状腺激素脱碘酶家族(thyroid hormone deiodinase families)等[13]。硒蛋白在生物体内有抗氧化、清除体内自由基的功能,硒的缺乏将会使癌症、病毒感染以及男性不育的几率增加[14-16]。硒的严重缺乏将会使动物患上白肌病,使人类患上克山病[17-18]。
农作物主要吸收转化土壤中的硒酸盐(selenate)和亚硒酸盐(selenite),为人及哺乳动物提供硒。不同植物的硒累积量具有很大差异,有的甚至可达到1 000 倍[19],这可能是不同植物的硒转运蛋白的选择性及转运能力不同造成[20-21]。
在植物体内,硒酸盐主要是通过硫酸盐转运蛋白进行吸收。而亚硒酸盐的吸收比较复杂,有多种因素[22-27]可影响其在植物体内的累积。最近研究发现,多种膜转运蛋白参与了亚硒酸盐的吸收累积,如硅转运蛋白[28]、磷酸盐转运蛋白[23,29]和单羧酸转运蛋白[30]等。本文主要对近几年植物硒转运蛋白的研究结果进行总结,并对未来的研究方向进行展望。
1 植物对硒的吸收
硒是非金属元素,有4 种同素异形体:灰色单质硒、红色单质硒、黑色单质硒和无定形硒,其中,黑色单质硒最为稳定。在自然界中,硒主要以-2,0,+4,+6 等不同价态存在于有机和无机化合物中,其中,无机硒化合物主要是硒酸盐和亚硒酸盐,而有机硒形态较多,但参与蛋白质合成的主要是硒代半胱氨酸(selenocysteine,SeCys)和硒代甲硫氨酸(selenomethionine,SeMet)。
1.1 硒酸盐和亚硒酸盐在环境中的分布
在不同土壤条件下,硒酸盐和亚硒酸盐的分布不同。在氧气比较充足、土壤比较干燥的碱性环境((pe+pH)>15)中,硒酸盐的含量较多;在潮湿、缺氧的酸性到中性土壤(7.5<(pe+pH)<15)中,亚硒酸盐的含量较多;在其他还原性较强的土壤((pe+pH)<7.5)中,硒化物(selenide)占主要地位[26,31]。植物吸收少量的有机硒化物[32],主要是通过不同的吸收机制吸收土壤中的硒酸盐和亚硒酸盐。
1.2 硒酸盐转运蛋白
硒和硫是同一主族元素,硒酸盐和硫酸盐物理化学性质相似[33],高浓度硫酸盐在一定程度上抑制硒酸盐的吸收[34],硫酸盐转运蛋白的突变可影响细胞对硒酸盐的耐受能力[35-36],表明硒酸盐可通过硫酸盐转运蛋白进入细胞内。
根据硫酸盐转运蛋白编码序列的同源性、动力学特性及组织和细胞定位的不同,硫酸盐转运蛋白可分为5 大类[37]。第I 类是高亲和性的硫酸盐转运蛋白(硫酸盐Km为1.5~10 μmol/L),如在拟南芥根部大量表达的AtSultr1;2 和AtSultr1;1[38],通常在韧皮部表达的AtSultr1;3[39]。第II类大部分是低亲和性的硫酸盐转运蛋白(硫酸盐Km值大于100 μmol/L),如在根、叶微管组织中表达较多的AtSultr2;1,根韧皮部及叶维管束鞘细胞表达的AtSultr2;2[40-41]。第III 类在种皮、胚中表达,且其表达不受培养介质中硫酸盐含量的影响[40]。第IV 类参与调节胞内硫酸盐浓度[42]。推测第V 类可能参与液泡硫酸盐的转运[37],但尚未得到证实。
第I 类硫酸盐转运蛋白对硫酸盐的高亲和性、在根系中大量表达以及受硫酸盐的表达调控[37],表明其在植物吸收硫酸盐的过程中具有重要作用。为研究此类转运蛋白是否也在硒酸盐吸收过程中发挥同样的功能,利用不同的拟南芥耐硒突变体sel1-11,sel1-12,sel1-13,sel1-14 发现第I 类硫酸盐转运蛋白AtSultr1;2 编码区的突变或者启动子突变均可提高其耐硒能力[36,43-44]。
AtSultr1;1 与AtSultr1;2 在吸收硫酸盐过程中具有相同的功能,但AtSultr1;2 突变影响硒酸盐吸收能力,而AtSultr1;1 突变不影响其吸收[36],说明AtSultr1;2 为硒酸盐转运蛋白。紫云英属高累积硒植物,茎部的硒含量可达干质量的0.6%,比非累积植物高100~1 000 倍[19],这可能是由于不同植物的硒酸盐转运蛋白的选择性及转运能力不同造成的[20]。
1.3 亚硒酸盐转运蛋白
亚硒酸为弱酸,其pKa1和pKa2分别为2.57和6.60[28]。亚硒酸盐在pH 值低于2.5 的环境中主要为H2SeO3/SeO2;在pH 值为2.5~6.6 的环境中主要为HSeO3-,在pH 值大于6.6 的环境中主要为SeO32-[28]。因其解离状况的不同,植物根系细胞对不同形态的+4 价态硒化合物的吸收能力可能不同。研究表明,水稻幼苗根在不同pH 值条件下对亚硒酸盐可能具有不同的吸收机制[26]。结果显示,pH 值为3.0,8.0 时,DNP 和低温(4 ℃)对水稻根系吸收硒的能力仅有较小程度抑制;而pH 值为5.0 时,DNP 和低温能够抑制大部分亚硒酸盐的吸收。进一步分析表明,在pH 值为3.0的条件下,亚硒酸盐可能主要是通过水孔蛋白被动进入根细胞;而在pH 值为5.0 的条件下,主要以主动方式进入根细胞[26]。
最近,Zhao 等[28]发现,属于水孔蛋白家族的水稻硅转运蛋白Lsi1(OsNIP2;1)可作为亚硒酸盐转运蛋白发挥功能(图1)。lsi1 水稻突变株与野生株相比,地上部分和木质部汁液中硒浓度明显降低,而硫酸盐的吸收不受影响;lsi2 突变株与野生株相比,亚硒酸盐的吸收能力几乎不受影响,表明水稻硅转运蛋白Lsi1 参与了亚硒酸盐的吸收,而Lsi2 没有参与此过程[28]。为了明确Lsi1对亚硒酸盐的运转活性,Zhao 等[28]在酵母中表达lsi1 时发现,在偏酸性(pH 值为3.5,5.5)条件下,Lsi1 可促进亚硒酸盐的吸收;而在偏碱性(pH 值为7.5)条件下,Lsi1 对亚硒酸盐的吸收能力没有明显变化。这可能是由于pH 值影响了亚硒酸盐的解离[28],在偏酸性条件下,主要是不带电荷的H2SeO3/SeO2可通过水孔蛋白进出细胞。
目前,尚没有在植物中发现HSeO3-转运蛋白。因为HSeO3-与单羧酸结构类似,McDermott 等[30]认为,亚硒酸盐可通过单羧酸转运蛋白Jen1p 进行吸收,且溶液pH 值会影响其转运活性,在pH值为4.5~7.5 范围内,随着pH 值降低,转运活性会提高;且质子梯度解偶联剂极大地抑制其转运活性[30],说明此转运过程为质子梯度推动的或者是ATP 依赖的主动转运过程。此结果与氰化物抑制单羧酸转运蛋白活性,抑制细胞对乳酸、丙酮酸等的吸收[45-46],以及解偶联剂和低温抑制水稻根细胞在pH 值为5.0 时对亚硒酸的吸收[26]是一致的。Lazard 等[29]发现,酵母磷酸盐转运蛋白在pH 值为6.0 时具有亚硒酸盐转运活性。磷酸的Ka1,Ka2,Ka3分别为7.25×10-3,6.31×10-8和3.98×10-13,在pH 值为6.0 时磷酸盐主要形态为H2PO4-,而亚硒酸盐主要形态为HSeO3-。我们推测因HSeO3-与H2PO4-结构类似,导致在此pH 值条件下,亚硒酸盐可通过磷酸盐转运蛋白进行转运。Li 等[23]也发现,植物磷酸盐转运蛋白参与了亚硒酸盐的吸收。
在偏碱性条件下,亚硒酸盐的主要形式为SeO32-[28],水稻的亚硒酸盐吸收量较酸性条件下吸收量少很多[26,28],导致此现象的原因可能为植物对SeO32-吸收能力低或者在此条件下外排亚硒酸盐能力提高。酵母亚硒酸盐外排蛋白SSU1在酵母的耐硒过程中[47]具有重要的功能。植物体内是否具有同样功能的蛋白尚没有报道。
亚硒酸盐吸收途径的多样性导致植物对亚硒酸盐的吸收量受到多种因素的影响,如呼吸抑制剂、代谢抑制剂、硝酸盐、磷素营养、pH 值及谷胱甘肽均可影响植物对亚硒酸盐的吸收[22-27]。由图1 可知,我们推测硒的跨膜转运是由多种转运蛋白参与的,除硒酸盐转运主要通过硫酸盐转运蛋白进行外,亚硒酸盐基于其在不同酸碱度情况下的解离形式,推测分别利用水孔蛋白类通道转运H2SeO3/SeO2,酵母磷酸盐和单羧酸转运蛋白转运HSeO3-,植物体内是否存在SeO32-转运蛋白以及亚硒酸盐外排蛋白尚不清楚。
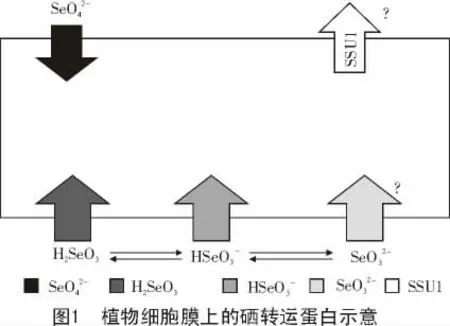
目前,大部分硒的吸收转运研究主要集中在植株水平上,为进一步分析不同转运蛋白吸收硒能力的差异,建立硒酸盐及亚硒酸盐转运蛋白的标准鉴定方法,进而分析不同转运蛋白的硒通透能力、亲和能力的差异,以及利用结构生物学手段分析其结构差异,为深入探讨不同植物耐硒能力差异的机理奠定基础。
2 硒在植物体内的同化代谢
植物从根部转运硒至地上部的转运效率取决于根从土壤中吸收硒的类型,硒酸盐被根系吸收后,可被运输到地上部分[23,25],在叶绿体内进行硒酸盐同化(图2)。而植物吸收的大部分亚硒酸盐累积于根部[25],木质部的亚硒酸盐含量很低[23,25]。这可能是由于亚硒酸盐在根部很快被转化为其他形式的硒化合物,如硒代甲硫氨酸(SeMet)和硒代甲硫氨酸水合物(selenomethionine Se-oxide hydrate,SeOMet)等,但大多数为非水溶性硒化合物[22-23]。
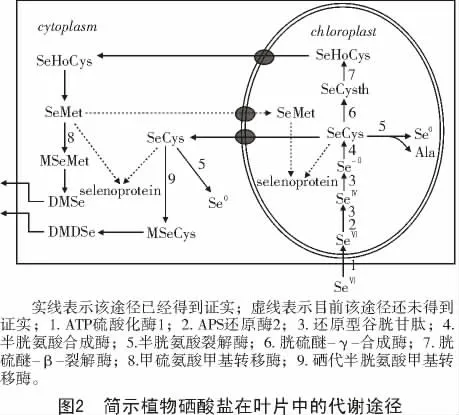
硒在植物体内的同化代谢过程已经被研究得较为清楚[21,48](图2),硒酸盐进入叶绿体后在ATP 硫酸化酶1(ATP sulfurylase 1,ATPS1)的催化下合成腺苷-5′-磷酰硒酸(Adenosine 5′-phosphoselenate,APSe)[49],APSe 可被APS 还原酶2(APS Reductase 2,APR2)还原形成亚硒酸盐[50-51]或者经GSH 的还原生成亚硒酸盐并进一步还原成Se2-[48],Se2-在半胱氨酸合成酶(Cys synthetase)的催化作用下合成SeCys。合成的SeCys 有多种代谢去路。SeCys 经半胱氨酸裂解酶(selenocysteine lyase,SL)裂解成Se0和丙氨酸;或者在胱硫醚-γ-合成酶(Cystathionine-γ-synthase)的催化下形成硒代胱硫醚(Secystathionine,SeCysth),后被胱硫醚-β-裂解酶(Cystathionine-β-lyase)裂解为硒代高半胱氨酸(Sehomocysteine,SeHo-Cys);也有部分SeCys 进入胞液,在胞液中被SL裂解生成Se0,或者被硒代半胱氨酸甲基转移酶(SeCys methyltransferase,SMT)甲基化形成甲基硒代半胱氨酸(MSeCys),MSeCys 在相关酶的作用下进一步甲基化形成具挥发性的二甲基二硒化物(DMDSe)[52-54]。另外一种形成挥发性硒化物的途径为:叶绿体内的SeHoCys 进入胞液内形成硒代甲硫氨酸,硒代甲硫氨酸被甲硫氨酸甲基转移酶(MMT)甲基化形成甲基甲硫氨酸(MSeMet),MSeMet 进一步甲基化形成挥发性二甲基硒化物(DMSe)。在叶绿体和胞液中的硒代半胱氨酸和硒代甲硫氨酸在一定条件下可取代半胱氨酸和甲硫氨酸参与蛋白质的合成,使蛋白不能正确折叠[55-56],甚至阻止mRNA 的翻译[57],从而导致植物生长受到抑制[58]。
3 展望
硒是一种重要的生命元素,它不仅是动物、人类和微生物的必需元素[1-3],也是植物生长发育的有益元素[4-8]。自然环境中硒主要以硒酸盐和亚硒酸盐2 种氧化形态存在,植物和微生物可以吸收环境中的硒酸盐和亚硒酸盐,将其转化为有机硒被人类和动物所利用。
硒酸盐在植物体内的同化代谢过程已较为清楚,普遍认为,植物吸收硒酸盐主要是通过硫酸盐转运蛋白进行。利用突变体的研究结果表明,即使是同一类型的硫酸盐转运蛋白,其转运硒酸盐的能力也可能不同[36]。可借鉴离子通道的鉴定分析方法,建立硒酸盐转运蛋白的标准鉴定方法,为进一步分析不同硒转运蛋白的转运能力及特异性,比较同一物种不同的转运蛋白以及不同物种转运蛋白的转运特异性及能力差异等奠定基础。
某些植物水孔蛋白和磷酸盐转运蛋白参与了亚硒酸盐的吸收[23,26,59],植物的单羧酸转运蛋白是否也具有亚硒酸盐吸收转运活性尚不清楚。根据目前的结果,我们推测亚硒酸盐的吸收转运途径为3 种对应于H2SeO3/SeO2,HSeO3-和SeO32-的转运蛋白和一种外运途径。这种亚硒酸盐吸收途径的多样性,可能是影响细胞代谢、亚硒酸盐形态等多种因素[22-27]以及亚硒酸盐吸收量的原因。建立亚硒酸盐转运的标准鉴定方法,为进一步分析鉴定不同形态亚硒酸盐的吸收转运载体及其特性、鉴定植物体内的候选硒酸盐外排蛋白等工作提供条件。
综上所述,为深入研究分析植物硒转运蛋白,亟待解决的问题包括:(1)建立鉴定硒转运蛋白标准方法;(2)分析不同转运蛋白的特性(Km,Vm,特异性等);(3)克隆分析植物磷酸盐转运蛋白和单羧酸转运蛋白等的硒转运特性;(4)比较硒累积能力具有很大差异的植物同源硒转运蛋白的转运能力差异;(5)明确植物是否外排硒转运蛋白等。这些问题的阐明将有助于我们深入探讨植物的富硒、耐硒原理,为进一步通过培育富硒粮食作物满足人类对硒的需求,培育耐硒植物进行高硒环境的生物修复等奠定基础。
[1]Fu L H,Wang X F,Eyal Y,et al. A selenoprotein in the plant kingdom:Mass spectrometry confirms that an opal codon(UGA)encodes selenocysteine in Chlamydomonas reinhardtii gluththione peroxidase [J]. Journal of Biological Chemistry,2002,277(29):25983-25991.
[2]Birringer M,Pilawa S,Flohe L.Trends in selenium biochemistry[J].Natural Product Reports,2002,19(6):693-718.
[3] Obata T,Shiraiwa Y. A novel eukaryotic selenoprotein in the haptophyte alga Emiliania huxleyi [J]. Journal of Biological Chemistry,2005,280(18):18462-18468.
[4]樊文华,李莉.硒钴配施对番茄产量、番茄红素及硒、钴含量的影响[J].山西农业科学,2009,37(10):19-22.
[5]唐玉霞,王慧敏,吕英华,等.冬小麦硒素吸收积累特性及叶面喷硒效应的研究[J].华北农学报,2010,25(S1):204-207.
[6] 张鹏飞,张爱军,张建恒,等.叶面施硒对谷子硒富集及品质的影响[J].华北农学报,2010,25(4):235-238.
[7]周勋波,吴海燕,张惠君,等.喷施硒肥对大豆生长发育和生理生态参数的影响[J].华北农学报,2004,19(4):80-83.
[8]段曼莉,胡斌,梁东丽,等.4 种蔬菜对硒酸盐的吸收、富集与转运特征的研究 [J]. 农业环境科学学报,2011,30(3):422-428.
[9]Yang G Y,Wang S Z,Zhou R.Endemic selenium intoxication of humans in China[J].The American Journal of Clinical Nutrition,1982,37:872-881.
[10]Hamilton S J. Review of selenium toxicity in the aquatic food chain[J].Sci Total Environ,2004,326(1/3):1-31.
[11] Fordyce F,Guangdi Z,Green K,et al. Soil,grain and water chemistry in relation to human selenium-responsive diseases in Enshi District,China[J].Applied Geochemistry,2000,15(1):117-132.
[12]朱建明,左维,秦海波,等.恩施硒中毒区土壤高硒的成因:自然硒的证据[J].矿物学报,2008,28(4):397-400.
[13]Kryukov G V,Castellano S,Novoselov S V,et al.Characterization of mammalian selenoproteomes [J]. Science,2003,300:1439-1443.
[14] Ellis D R,Sors T G,Brunk D G,et al. Production of Se-methylselenocysteine in transgenic plants expressing selenocysteine methyltransferase[J].Bmc Plant Biology,2004,4:1-11.
[15]White P J,Broadley M R. Biofortification of crops with seven mineral elements often lacking in human diets—iron,zinc,copper,calcium,magnesium,selenium and iodine[J]. New Pytol,2009,182(1):49-84.
[16]Diwadkar-Navsariwala V,Prins G S,Swanson S M,et al. Selenoprotein deficiency accelerates prostate carcinogenesis in a transgenic model[J].Proc Natl Acad Sci USA,2006,103(21):8179-8184.
[17]Fairweather-Tait S J,Bao Y,Broadley M R,et al. Selenium in human health and disease [J].Antioxid Redox Signal,2011,14(7):1337-1383.
[18]Ishihara H,Kanda F,Matsushita T,et al.White muscle disease in humans:myopathy caused by selenium deficiency in anorexia nervosa under long term total parenteral nutrition [J]. J Neurol Neurosurg Psychiatry,1999,67(6):829-830.
[19]Pickering I J,Wright C,Bubner B,et al. Chemical form and distribution of selenium and sulfur in the selenium hyperaccumulator Astragalus bisulcatus [J]. Plant Physiol,2003,131(3):1460-1467.
[20]Galeas M L,Zhang L H,Freeman J L,et al. Seasonal fluctuations of selenium and sulfur accumulation in selenium hyperaccumulators and related nonaccumulators [J]. New Phytol,2007,173(3):517-525.
[21]Zhu Y G,Pilon-Smits E A,Zhao F J,et al.Selenium in higher plants:understanding mechanisms for biofortification and phytoremediation [J]. Trends in Plant Science,2009,14(8):436-442.
[22]Arvy M.Selenate and selenite uptake and translocation in bean plants(Phaseolus vulgaris)[J].Journal of Experimental Botany,1993,44(6):1083-1087.
[23]Li H F,McGrath S P,Zhao F J.Selenium uptake,translocation and speciation in wheat supplied with selenate or selenite[J].NewPhytologist,2008,178(1):92-102.
[24]Xu W F,Chen Q X,Shi W M. Effects of nitrate supply site on selenite uptake by rice roots [J]. J Agric Food Chem,2010,58(20):11075-11080.
[25]陈思杨,江荣风,李花粉.苗期小麦和水稻对硒酸盐/亚硒酸盐的吸收及转运机制 [J]. 环境科学,2011,32(1):284-289.
[26]张联合,李友军,苗艳芳,等.pH 值对水稻离体根系吸收亚硒酸盐生理机制的影响 [J]. 土壤学报,2010,47(3):523-528.
[27]张联合,郁飞燕,李友军,等.L-半胱氨酸对水稻根系吸收亚硒酸盐的影响[J].土壤学报,2010,47(5):1018-1022.
[28]Zhao X Q,Mitani N,Yamaji N,et al.Involvement of silicon influx transporter OsNIP2;1 in selenite uptake in rice [J]. Plant Physiol,2010,153(4):1871-1877.
[29]Lazard M,Blanquet S,Fisicaro P,et al. Uptake of selenite by Saccharomyces cerevisiae involves the high and low affinity orthophosphate transporters[J].Journal of Biological Chemistry,2010,285(42):32029-32037.
[30]McDermott J R,Rosen B P,Liu Z.Jen1p:a high affinity selenite transporter in yeast [J]. Molecular Biology of the Cell,2010,21(22):3934-3941.
[31]Elrashidi M,Adriano D,Workman S,et al.Chemical equilibria of selenium in soils:a theoretical development[J].Soil Science,1987,144:141-152.
[32]Abrams M,Shennan C,Zasoski R,et al.Selenomethionine uptake by wheat seedlings[J].Agron J,1990,82(1):127-131.
[33] Wessjohann L A,Schneider A,Abbas M,et al. Selenium in chemistry and biochemistry in comparison to sulfur [J]. Biol Chem,2007,388(10):997-1006.
[34]White P J,Bowen H C,Parmaguru P,et al.Interactions between selenium and sulphur nutrition in Arabidopsis thaliana [J]. J Exp Bot,2004,55:1927-1937.
[35]Cherest H,Davidian J C,Thomas D,et al.Molecular characterization of two high affinity sulfate transporters in Saccharomyces cerevisiae[J].Genetics,1997,145(3):627-635.
[36]Shibagaki N,Rose A,McDermott J P,et al. Selenate-resistant mutants of Arabidopsis thaliana identify Sultr1;2,a sulfate transporter required for efficient transport of sulfate into roots[J].Plant J,2002,29(4):475-486.
[37] Buchner P,Takahashi H,Hawkesford M J. Plant sulphate transporters:co-ordination of uptake,intracellular and longdistance transport[J].J Exp Bot,2004,55:1765-1773.
[38] Yoshimoto N,Takahashi H,Smith F W,et al. Two distinct high-affinity sulfate transporters with different inducibilities mediate uptake of sulfate in Arabidopsis roots [J]. Plant J,2002,29(4):465-473.
[39]Yoshimoto N,Inoue E,Saito K,et al.Phloem-localizing sulfate transporter,Sultr1;3,mediates re-distribution of sulfur from source to sink organs in Arabidopsis [J]. Plant Physiol,2003,131(4):1511-1517.
[40]Takahashi H,Watanabe-Takahashi A,Smith F W,et al. The roles of three functional sulphate transporters involved in uptake and translocation of sulphate in Arabidopsis thaliana[J].Plant J,2000,23(2):171-182.
[41] Takahashi H,Yamazaki M,Sasakura N,et al. Regulation of sulfur assimilation in higher plants:a sulfate transporter induced in sulfate-starved roots plays a central role in Arabidopsis thaliana [J]. Proc Natl Acad Sci USA,1997,94(20):11102-11107.
[42]Kataoka T,Watanabe-Takahashi A,Hayashi N,et al. Vacuolar sulfate transporters are essential determinants controlling internal distribution of sulfate in Arabidopsis [J]. Plant Cell,2004,16(10):2693-2704.
[43] Cram W J,Kok LJd,Stulen I,et al. Sulphur metabolism in higher plants:molecular,ecophysiological and nutritional aspects[M].Leiden:Backhuys Publishers,1997:217-219.
[44]El Kassis E,Cathala N,Rouached H,et al.Characterization of a selenate-resistant Arabidopsis mutant:Root growth as a potential target for selenate toxicity[J].Plant Physiol,2007,143(3):1231-1241.
[45]Garcia C K,Brown M S,Pathak R K,et al. cDNA cloning of MCT2,a second monocarboxylate transporter expressed in different cells than MCT1 [J]. Journal of Biological Chemistry,1995,270(4):1843-1849.
[46]Cheeti S,Warrier B K,Lee C H. The role of monocarboxylate transporters in uptake of lactic acid in HeLa cells [J]. Int J Pharm,2006,325(1/2):48-54.
[47]Park H,Bakalinsky A T.SSU1 mediates sulphite efflux in Saccharomyces cerevisiae[J].Yeast,2000,16(10):881-888.
[48]Terry N,Zayed A M,de Souza M P,et al. Selenium in higher plants[J].Annual Review of Plant Physiology and Plant Molecular Biology,2000,51:401-432.
[49]Pilon-Smits E,De Souza M,Hong G,et al.Selenium volatilization and accumulation by twenty aquatic plant species[J].J Environ Qual,1999,28(3):1011-1018.
[50] Loudet O,Saliba-Colombani V,Camilleri C,et al. Natural variation for sulfate content in Arabidopsis thaliana is highly controlled by APR2[J].Nat Genet,2007,39(7):896-900.
[51]Grant K,Carey N M,Mendoza M,et al.Adenosine 5'-phosphosulfate reductase (APR2)mutation in Arabidopsis implicates glutathione deficiency in selenate toxicity [J]. Biochem J,2011,438(2):325-335.
[52]Freeman J L,Zhang L H,Marcus M A,et al. Spatial imaging,speciation,and quantification of selenium in the hyperaccumulator plants Astragalus bisulcatus and Stanleya pinnata [J].Plant Physiol,2006,142(1):124-134.
[53]LeDuc D L,Tarun A S,Montes-Bayon M,et al.Overexpression of selenocysteine methyltransferase in Arabidopsis and Indian mustard increases selenium tolerance and accumulation [J].Plant Physiol,2004,135(1):377-383.
[54]Leduc D L,AbdelSamie M,Montes-Bayon M,et al. Overexpressing both ATP sulfurylase and selenocysteine methyltransferase enhances selenium phytoremediation traits in Indian mustard[J].Environmental Pollution,2006,144(1):70-76.
[55]Birringer M,Pilawa S,Flohe L.Tends in selenium biochemistry[J].Natural Product Reports,2002,19:693-781.
[56]Brown T,Shrift A. Selenium: toxicity and tolerance in higher plants[J].Biological Reviews,1982,57(1):59-84.
[57]Eustice D C,Kull F J,Shrift A.Selenium toxicity:aminoacylation and peptide bond formation with selenomethionine[J].Plant Physiol,1981,67(5):1054-1058.
[58]段曼莉,付冬冬,王松山,等.亚硒酸盐对四种蔬菜生长、吸收及转运硒的影响 [J]. 环境科学学报,2011,31(3):658-665.
[59] Hu H,Wang M,Zhan X,et al. Effect of different selenium sources on productive performance,serum and milk Se concentrations,and antioxidant status of sows [J].Biol Trace Elem Res,2011,142(3):471-480.
猜你喜欢
中国生物化学与分子生物学报(2022年7期)2022-09-07
中国药科大学学报(2021年6期)2021-12-31
云南化工(2021年5期)2021-12-21
国际放射医学核医学杂志(2020年2期)2020-05-30
四川冶金(2019年5期)2019-12-23
食品科学(2016年3期)2016-11-11
中国煤炭(2016年1期)2016-05-17
中国资源综合利用(2016年1期)2016-02-03
东北电力大学学报(2015年4期)2015-11-13
应用化工(2014年8期)2014-08-08
