
局部场电位γ频带能量对朝向调谐特性研究
2012-12-03 01:23:04尚志刚冯平艳刘新玉牛晓可
郑州大学学报(工学版) 2012年6期
尚志刚,冯平艳,刘新玉,牛晓可,万 红
(郑州大学电气工程学院,河南郑州450001)
0 引言
朝向调谐特性是初级视觉皮层(Primary Visual Cortex,又称V1区)神经元感受野的主要特性之一[1].朝向调谐特性的传统研究方法是通过使用神经元spike发放率作为其朝向调谐指标.然而,由于在spike检测和分类过程中经常会发生假阳性和假阴性等问题[2],严重影响了后续分析的结果.
局部场电位(Local Field Potential,LFP)是电极尖端附近神经元响应信号的总和,频带主要集中在1~200 Hz[3].LFP低频部分主要反映状态信息,高频部分尤其是γ频带(30~90 Hz),主要反映了与视觉刺激相关的信息.Juergens等[4]在研究初级视觉皮层局部场电位信号和EEG信号的关系时发现γ频带的能量反映了视觉刺激的信息.Frien等[5]研究指出LFPγ频带的功率会随着刺激朝向的改变而产生不同的调制作用;Siegel和König[6]研究也发现γ频带的能量会随着朝向的改变而改变;Henrie等[7]用运动光栅刺激研究发现局部场电位γ频带的能量在不同的朝向下有选择性的加强.但文献中没有给出两种信号对朝向调谐特性研究的定量比较.
以大鼠为研究对象,将微电极阵列植入大鼠V1区,以不同朝向的全屏光栅为视觉刺激,同时记录LFP和spike信号,研究γ频带能量对光栅朝向的调制作用.结果表明:LFPγ频带能量和spike发放率获得的神经元偏爱朝向一致率达到了67.4%,朝向调谐一致率达到了87.2%.并且仿真研究结果发现,基于LFPγ频带能量的朝向调谐指数比spike发放率具有更强的抗噪能力.
1 原理与方法
1.1 朝向调谐指数的计算
不同朝向光栅刺激LFPγ频带能量朝向调谐指数 ρLFP
[8]计算如下:

若ρLFP>0.7,则该神经元具有偏爱朝向.其中Pp为最优朝向γ频带能量,Po为与最优朝向正交的两个朝向γ频带能量P1、P2的均值,即:Po=(P1+P2)/2.

不同朝向光栅刺激spike发放率朝向调谐指数ρs可类似计算:若ρs>0.7,则该神经元具有偏爱朝向.其中Pp为最优朝向spike发放率;Po为与最优朝向正交的两个朝向 spike发放率 P3、P4的均值,即:Po=(P3+P4)/2.
偏爱朝向一致率与朝向调谐一致率定义为:设总神经元个数为M,其中两种信号都有偏爱朝向并且偏爱朝向一致的神经元个数为N1,两种都没有偏爱朝向的神经元个数为N2,偏爱朝向一致率为λ1=N1/M,朝向调谐一致率为λ2=(N1+N2)/M.
1.2 LFPγ频带的提取和功率谱估计
首先对原始数据使用通带为0~250 Hz的二阶Butterworth带通滤波器对信号进行滤波,然后对滤波后的信号重采样,得到采样频率为2 kHz的局部场电位信号.
由于LFP为非平稳非高斯信号,在检测的过程中往往会出现一些短时瞬态脉冲,这些瞬态脉冲信号可能包含有刺激的信息.传统的瞬态脉冲检测方法中,匹配滤波[9]是最常用的一种,但是LFP非平稳性极强,匹配模板需要不断更新.因此,采用小波变换的方法,用逐渐精细的时间分辨率逼近信号的快变部分,用逐渐精细的频率分辨率逼近信号的慢变部分.
小波基函数ψ(t)不具唯一性,不同的小波基函数分析得到的结果相差甚远.笔者基于LFP信号的特点,选择与LFP信号相似的‘db5’作为小波的基函数.
小波分解公式为:
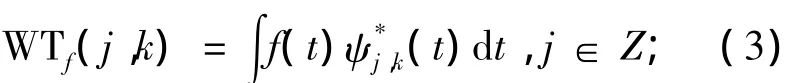
小波重构公式为:

其中,小波基函数为:

式中:a0>0;j,k∈Z;f(t)为原信号;a-j0为尺度因子,kτ0为平移因子.
LFP信号采样频率为2 kHz,为了精确获得γ频带信号,按照式(3)的小波分解算法对原始信号进行5层小波分解,经过5层分解后,选择其中62.5~125 Hz频带再分解后获得62.5~93.75 Hz频带,利用此频带与第一次分解得到的31.25~62.5 Hz频带进行重建,得到γ频带信号(30~90 Hz),重构出的γ频带信号如图1所示.小波重构出的γ频带较好地抑制了噪声并且保留了响应的瞬态特征.
笔者选用Welch功率谱估计对LFP进行功率估计.Welch功率谱估计的关键是选取合适的窗和窗长以适应不同分辨率的要求.因此笔者选择分辨率和方差水平都较好的Hamming窗进行谱估计.选用的Hamming窗分辨率为8 Hz,局部场电位频带信号带宽约为60 Hz,采样频率2 kHz.对比研究发现:分辨率8 Hz较好地抑制了噪声,并且可以准确地提取出频带的能量,其对应的窗长N=500,其功率谱估计如图2所示.由图2可以看出,该分辨率下谱估计方差和噪声水平都较好,频带能量主要集中在35~60 Hz频带内.

2 实验分析
2.1 实验数据采集
实验采用年龄11周左右,体重200~250 g的成年雄性Long Evans(LE)大鼠.术前实验动物用水合氯醛腹腔注射麻醉后,固定于Stoelting脑立体定位仪上,参考大鼠脑立体定位图谱在V1区植入2×8铂铱合金Microprobes微电极丝阵列.植入电极1.5 h后,对大鼠进行12个朝向(0~330°,间隔30°)的全屏光栅视觉刺激,并用 CerebusTM数据采集系统记录大鼠V1区初级视觉皮层信号.
2.2 LFPγ频带能量分析
为了研究γ频带能量是否能反应刺激的信息,笔者首先分析了有光栅刺激和无光栅刺激γ频带的能量分布.图3为其中一个通道的分析结果.由图3可以看出:γ频带能量主要集中在35~60 Hz,并且有刺激时γ频带能量比无刺激时有较为明显的变化,说明了γ频带的能量编码了刺激的信息.

图3 有刺激和无刺激γ频带能量图Fig.3 Power of gamma band with simulation and without simulation
为了研究γ频带的能量对于光栅朝向的调制作用,笔者采用最佳参数光栅刺激(时间频率4 Hz,空间频率0.2 cpd),随机变换其朝向,其中一个通道偏爱朝向和正交朝向的分析结果如图4所示.由图4可知,不同的朝向引起的神经元响应不同,其中偏爱朝向刺激前后γ频带能量差变化最为显著.

图4 偏爱朝向和正交朝向刺激前后γ频带能量差Fig.4 Power increase of the preferred and orthogonal orientation
2.3 spike的提取和分类
对原始数据使用通带0.25~3 kHz的二阶带通Butterworth滤波器滤波,然后进行阈值检测,阈值水平设定为信号标准差的5倍[10].利用主成分分析(Principal Component Analysis,PCA)结合 kmeans聚类进行spike的分类提取[11].
某通道第一和第二主成分利用k-means聚类后如图5(b)所示.锋电位被分为明显的两类,选用发放较多的一类(图5(c))做研究,其朝向调谐曲线如图5(d)所示.由图可以看出,该神经元对90°光栅表现出较为明显的朝向选择性.
2.4 LFPγ频带能量和spike的朝向调谐分析
2.4.1 LFP与spike朝向调谐对比
分别用两种信号研究了172个神经元的朝向调谐特性.其中两种信号有150个神经元(116个有偏爱朝向,34个无偏爱朝向)的分析结果保持一致,有偏爱朝向的一致率达到了67.4%,总体一致率达到了87.2%.这说明γ频带能量可以做为研究朝向选择性的有效指标,其中一个神经元的对比结果如图6所示.

图6为某一通道两种方法的朝向调谐曲线图.图6(a)为基于LFPγ频带能量的朝向调谐曲线,朝向调谐指数ρLFP=0.710 9;图6(b)为基于spike发放率的朝向调谐曲线,朝向调谐指数ρs=0.704 1.可以看出,该神经元LFP频带能量在研究朝向调谐特性时能达到和spike发放率一样的效果.
2.4.2 不同信噪比下实验数据分析
为研究利用LFP和spike两种信号分析朝向调谐特性时的抗噪能力,笔者研究了不同信噪比下两种信号的分析结果.实验数据由实测信号加白噪声组成.从原始信号中提取出spike信号和LFPγ频带信号,重构成纯净的神经放电信号,如图7所示.

图7 仿真数据的建立Fig.7 The simulation data generation
将重构的原始纯净信号加入不同SNR的高斯白噪声[12],分析不同信噪比下两种信号的朝向调谐特性.针对LFP频带能量和spike发放率,计算在不同信噪比时两种信号的朝向调谐指数和偏爱朝向.两种方法下偏爱朝向以及调谐指数的分析结果如表1所示.

表1 不同信噪比下LFPγ频带能量与spike发放率的比较Tab.1 The comparison between LFP Gamma band power and spike under different SNR
由表1可以看出:随着信噪比的降低,两种算法的朝向调谐指数都有所降低,其中基于spike发放率的朝向调谐指数降低更快.当信噪比在0 dB以下时,基于spike发放率的偏爱朝向出现了改变.这说明,与spike发放率相比,LFPγ频带能量在计算神经元感受野朝向调谐特性方面具有更强的抗噪能力.
3 结论
利用LFP信号γ频带能量分析了V1区神经元感受野的朝向调谐特性.用小波变换结合Welch功率谱估计提取并计算了局部场电位γ频带在光栅朝向刺激下的能量分布,并对比分析了LFP和spike两种信号下的研究结果.
研究表明,与传统基于spike发放率的朝向调谐特性计算方法相比,基于LFPγ频带能量计算方法具有更强的抗噪能力,在低信噪比的情况下也能有效地估计出神经元感受野的朝向调谐特性.因此,LFPγ频带能量在低信噪比的情况下,研究V1区神经元感受野的朝向调谐特性方面比spike发放率更有优势,为研究大鼠V1区神经元感受野的朝向调谐特性提供了另一种可行的方法.
[1] MELIZA C D,DAN Y.Receptive field modification in rat visual cortex induced by paired visual stimulation and single cell spiking[J].Neuron,2006,49:183-189.
[2] HILL D N,MEHTA SB,KLEINFELD D.Quality metrics to accompany spike sorting of extracellular signals[J].The Journal of Neuroscience,2011,31(24):8699-8705.
[3] GAWNE T J.The local and nonlocal components of the local field potential in awake primate visual cortex[J].Journal of Computational Neuroscience,2010,29:615-623.
[4] JUERGENS E,GUETTLER A,ECKHORN R.Visual stimulation elicits locked and induced gamma oscillations in monkey intracortical and EEG potentials,but not in human EEG [J].Experimental Brain Research,1999,129:247-259.
[5] FRIEN A,ECKHOMR,WOELBEMT,et al.Fast oscillations display sharper orientation tuning than slower components of the same recordings in striate cortex of the awake monkey[J].European Journal of Neuroscience,2000,12:1453-1465.
[6] SIEGE L M,KNIG P.Frequency distribution of stimulus dependent synchronization in area 18 of the awake cat[J].The Journal of Neuroscience,2003,23:4251-4260.
[7] HENNE J A,SHAPLEY R.LFP power spectra in V1 cortex:the graded effect of stimulus contrast[J].
Journal of Neurophysiology,2005,94(1):479-490.[8] CARANDINIA M,FERSTE D.Membrane potential and
firing rate in cat primary visual cortex[J].The Journal of Neuroscience,2000,20(1):470-484.
[9] 万红,郜丽赛,牛晓可.基于匹配小波变换的初级视皮层神经元锋电位分类[J].郑州大学学报:工学版,2011,32(3):90-94.
[10] VARGAS-IRWIN C,DONOGHUE J P.Automated spike sorting using density grid contour clustering and subtractive waveform decomposition[J].Journal of Neuroscience Methods,2007,164(1):1-18.
[11] LEWICKI MS.A review of methods for spike sorting:the detection and classification of neural action potentials[J].Computation in Neural Systems,1998(9):53-78.
[12] 王静,封洲燕.多通道神经元锋电位检测和分类的新方法[J].生物化学与生物物理进展,2009,36(5):641-647.
猜你喜欢
辽河(2022年1期)2022-02-14 21:48:35
辽河(2022年1期)2022-02-14 05:15:04
电脑知识与技术·经验技巧(2020年5期)2020-06-22 13:19:24
航天电子对抗(2019年4期)2019-06-02 08:22:44
读者·校园版(2018年24期)2018-12-04 03:04:44
电子测试(2017年15期)2017-12-18 07:18:51
高师理科学刊(2016年8期)2016-06-15 20:27:46
湖南城市学院学报(自然科学版)(2016年4期)2016-02-27 14:02:41
电测与仪表(2015年12期)2015-04-09 11:44:50
上海制造(2013年11期)2014-03-26 08:02:06
