
miR-214表达载体构建及其对靶基因β-catenin的调控
2012-11-26 12:39林述琨王耀鹏温吉海
药品评价 2012年36期
林述琨 王耀鹏 温吉海
1.大连普兰店市中心医院病理科; 2.普兰店市中心医院肺外科
MicroRNAs(miRNA)是一类长约19~25bp的单链非编码小分子RNA,能够与靶 基因mRNA的3UTR以不完全互补的方式结合,在转录后水平上引起靶基因mRNAs发生降解或翻译抑制[1,2]。近年研究发现miRNAs在多种人类肿瘤的发生发展中具有类似癌基因或抑癌基因的作用[3,4]。
研究报道miR-214可能参与多种肿瘤的发生和发展过程[5-8];本研究发现miR-214在肺癌组织中表达明显低于癌旁肺组织,同时我们构建了miR-214的真核表达载体,并通过双荧光素酶报告分析和Western blot方法探讨miR-214对靶基因的调控作用。从而为研 究miR-214在肺癌中作用机制提供了基础。
1材料和方法
1.1细胞培养和转染人肺腺癌细胞株A549,用含有10%新生牛血清,100U/ml的青霉素和100U/ml的链霉素的DMEM培养液在37℃、5%CO2饱和湿度条件下培养。当细胞生长至对数生长期后,按照lipo2000(Invitrogen公司)说明书的方法进行转染。细胞转染48h后提取细胞总RNA和蛋白进行后续相关实验。
1.2 Real-time PCR用Trizol(Invitrogen公司)提取细胞总RNA后反转录为cDNA(Takara)。miR-214的反转录引物为5′-GTC GTA TCC AGT GCA GGG TCC GAG GTA TTC GCA CTG GAT ACG ACA CTG CC-3′;real-time PCR引物序列:上游引物5′- GGA CAG CAG GCA CAG ACA -3′,下游引物5′- CAG TGC AGG GTC CGA GGT -3′。以U6为内参,反转录引物序列为5′-GTC GTA TCC AGT GCA GGG TCC GAG GTA TTC GCA CTG GAT ACG ACA AAA TAT GGA AC-3′;real-time PCR引物序列:上游引物5′-TGC GGG TGC TCC GCT TCG GCA GC-3′,下游引物5′-CAG TGC AGG GTC CGA GGT-3′。使用SYBR Green(Takara)染料法进行反应,95℃预变性10 min,95℃变性5s,60℃退火34s,74℃荧光检测3s,共40个循环。以2 值比较两者之间的差异。每组重复设置3个复孔。
1.3 miR-214质粒的构建根据 miR-214前体在GenBank中的序列,设计PCR引物,引物序列为:Forward:5'- AAGCTTCTGTTACGCAAATTATCCATGTT -3',Reverse:5'- CTCGAGAATGGGTTTTATTATATTTCATA-3',目的片段亚克隆到pCDNA3.1(+)上,经Real-time PCR验证重组质粒pCDNA3.1(+)-miR-214 能够过表达成熟的miR-214。
1.4 双荧光素酶报告基因检测应用生物信息学预测软件对miR-214进行靶基因预测,选β-catenin为候选靶基因,PCR扩增包括miR-214结合位点在内的β-catenin 3'UTR区,引物序列为:Forward:5'-AAT TGTAATCTGAATAAAGTGTAACAAT-3',Reverse:5'-ATGAATTAAAAGTTTAATTCTGAACCAT-3'(引入SacI和XhoI 酶切位点,), 目的片段亚克隆到 pMIR载体萤火虫荧光素基因的下游,重组载体命名为pMIR-β-catenin。对pMIR-β-catenin重组质粒“种子区”进行定点突变,突变后的质粒命名为pMIR-βcatenin-Mut。所有构建的表达质粒均经双酶切和测序鉴定。A549细胞在24孔板中培养,细胞融合度为80%左右时进行转染,转染分组为:pCDNA 3.1(+)-miR- 214+pMIR-β-catenin组、pCDNA3.1(+)+pMIR-β-catenin、pCDNA3.1(+)-miR- 214+pMIR-β-catenin-Mut组,每组设3个复孔。转染后继续培养48h,加入裂解液(100μl/孔),离心并收集上清至96孔板中,每孔中加入40μl 萤火虫荧光素酶底物,混匀10s后检测荧光强度;然后再加入40μl海肾荧光素酶底物,混匀10s后检测荧光强度。将萤火虫荧光素强度值/海肾荧光素强度值进行标准化校正。
1.5 Western blot检测β-catenin蛋白表达水平,用RIPA细胞裂解液(含有0.1%的PMSF)冰上裂解细胞20min,收集于1.5ml EP管中,离心后上清液转移至无菌EP管中,BCA法测定蛋白浓度。取等量蛋白在SDS-PAGE中电泳,转移到PVDF膜上;5%脱脂奶粉4℃过夜封闭,用β-catenin的兔抗人单克隆一抗(1:1000)室温下孵育2 h(cell signaling 9581),再用过氧化物酶标记的羊抗兔二抗(1:5000)室温下孵育2h,以β-actin为内参,用Bandscan 5.0软件分析蛋白条带总灰度值,检测表达差异。
1.6统计学方法应用SPSS 13.0统计学软件进行统计学分析。采用t检验进行两样本间比较,P<0.05为差异有统计学意义。
2 结果
2.1 miR-214在肺癌组织中的表达情况为了验证miR-214在肺癌中的表达情况,我们应用real-time PCR检测了5对肺癌组织及相应癌旁肺组织中miR-214的表达。结果显示,肺癌组织较癌旁肺组织miR-214明显低表达,见图1。
2.2 生物信息学预测β-catenin是miR-214的靶基因应用生物信息学预测软件 TargetScan对miR-214的靶基因进行预测,分析显示在β-catenin的3'UTR 区有7 个碱基与β-catenin 的“种子区”完全互补配对(见图2),因此生物信息学预测认为β-catenin可能是miR-214的靶基因。

图1 Real-time PCR方法检测miR-214的表达

图2 生物信息学软件预测miR-214的靶基因
2.3 双荧光素酶报告基因分析miR-214对β-catenin的调控作用将重组表达载体pCDNA3.1-miR-214和融合β-catenin 3'UTR 的表达质粒pMIR-β-catenin共转染A549 细胞后荧光素酶活性表达受到明显的抑制,而转染空载体pCDNA 3.1 和融合基因β-catenin'UTR的表达质粒pMIR-Aβ-catenin的荧光素酶活性无明显的变化;同样,转染pCDNA3.1-miR-214和融合β-catenin 3'UTR“种 子 区 ”突 变 的 表 达 质 粒pMIR-β-catenin-Mut 共转染A549细胞后荧光素酶活性没有受到抑制,与对照组转染空载体pCDNA3.1和融合基因β-catenin 3'UTR 的表达质粒的荧光素酶活性比较差异无统计学意义(见图3)。结果表明miR-214 能够对人β-catenin 3'UTR 区起到抑制性的调控作用。
2.4 Western blot检测miR-214抑制内源β-catenin蛋白表达 pCDNA 3.1-miR-214转染组中β-catenin蛋白表达与空白对照组(mock)和空载体转染组(pCDNA 3.1)中β-catenin蛋白表达差异明显,β-catenin在pCDNA 3.1-miR-214转染组中表达明显下调。结果表明miR-214能够在翻译水平上抑制β-catenin蛋白表达。
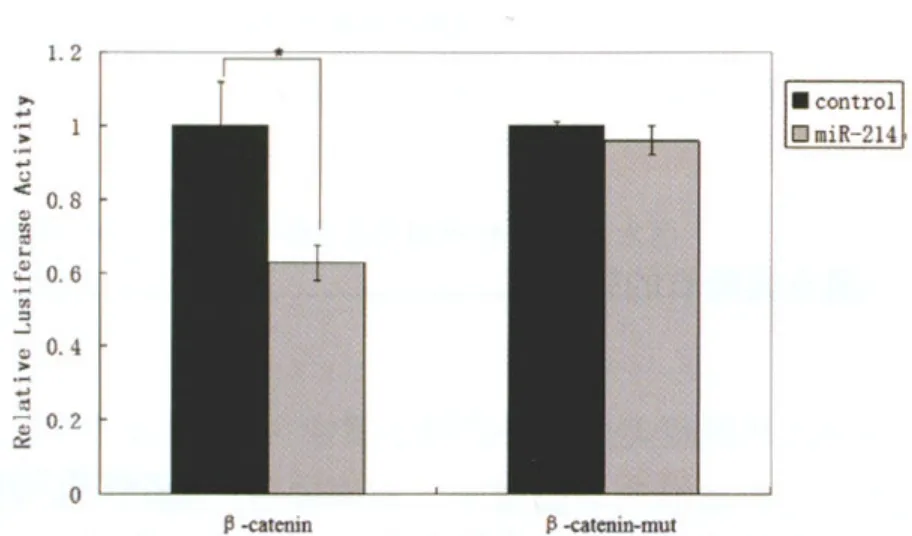
图3 双荧光素酶报告基因分析miR-214对β-catenin的调控

图4 Western blot检测β-catenin蛋白表达。1-空白对照组(mock);2-空载体转染组(pCDNA 3.1);3-pCDNA 3.1-miR-214转染组
3 讨论
目前,miRNA是现今肿瘤学的研究热点之一,miRNA可以与多种靶基因结合在肿瘤发生发展过程中可起到癌基因或抑癌基因作用[3,4]。研究报道miR-214可以抑制肝癌、宫颈癌和乳腺癌的侵袭和转移[5-8],起到抑癌基因作用。而目前miR-214在肺癌中的作用未见报道,现就miR-214在肺癌中的表达及靶基因进行了实验研究。
我们首先检测了肺癌和癌旁组织中miR-214的表达水平,发现miR-214在肺癌组织中表达明显下调,miR-214可能作为抑癌基因参与肺癌的发生。进一步我们通过生物信息学软件预测β-catenin可能是miR-214的靶基因,本研究首先通过双荧光素酶报告基因证实miR-214对的β-catenin的3'UTR 区具有作用的基础上,进一步在A549细胞中通过Western blot 方法证实了miR-214 对β-catenin的表达具有抑制作用。已有研究表明β-catenin的表达与肺癌的发生、转移相关[9,10]。因此,本研究对深入探讨miR-214 对β-catenin表达抑制作用的作用机理,为进一步研究miR-214在肺癌中的作用提供了思路和治疗的靶点。
[1] ZHANG H,LI Y,LAI M.The microRNA network and tumor metastasis[J].Oncogene,2010,29(7):937-948.
[2] OCANA OH,NIETO MA.A new regulatory loop in cancer-cell invasion[J].EMBO Rep,2008,9(6):521-522.
[3] SANTARPIA L,NICOLOSO M,CALIN G A.MicroRNAs:a complex regulatory network drives the acquisition of malignant cell phenotype[J].Endocr Relat Cancer,2010,17(1):F51-75.
[4] VOORHOEVE PM.MicroRNAs:Oncogenes,tumor suppressors or master regulators of cancer heterogeneity?[J].Biochim Biophys Acta,2010,1805(1):72-86.
[5] C.C.Wong,C.M.Wong,E.K.Tung,et al.The microRNA miR-139 suppresses metastasis and progression of hepatocellular carcinoma by down-regulating Rho-kinase 2[J].Gastroenterolo gy,2011,140(1):322–331.
[6] Wang Y,Lee AT,Ma JZ,et al.Profiling microRNA expression in hepatocellular carcinoma reveals microRNA-224 up-regulation and apoptosis inhibitor-5 as a microRNA-224-specific target[J].J Biol Chem,2008,283(19):13205–13215.
[7] Yang Z,Chen S,Luan X,et al.MicroRNA-214 is aberrantly expressed in cervical cancers and inhibits the growth of HeLa cells[J].IUBMB Life,2009,61(11):1075-1082.
[8] Derfoul A,Juan AH,Difilippantonio MJ,et al.Decreased microRNA-214 levels in breast cancer cells coincides with increased cell proliferation,invasion and accumulation of the Polycomb Ezh2 methyltransferase[J].Carcinogenesis,2011,32 (11):1607–1614.
[9] Yang LH,Xu HT,Han Y,et al.Axin downregulates TCF-4 transcription via beta-catenin,but not p53,and inhibits the proliferation and invasion of lung cancer cells[J].Mol Cancer,2010,2;9:25-30.
[10] Chiu CG,Chan SK,Fang ZA,et al.Beta-catenin expression is prognostic of improved non-small cell lung cancer survival[J].Am J Surg,2012,203(5):654-659.
猜你喜欢
中国现代医生(2022年21期)2022-08-22
天津医科大学学报(2021年4期)2021-08-21
现代畜牧科技(2021年4期)2021-07-21
广东蚕业(2021年1期)2021-03-18
天津医科大学学报(2021年1期)2021-01-26
医学信息(2019年9期)2019-06-10
江苏农业科学(2019年23期)2019-03-03
医学信息(2016年29期)2016-11-28
科技创新导报(2014年28期)2014-12-01
食品工业科技(2014年13期)2014-03-11
