
太湖秀丽白虾繁殖生物学研究
2012-11-10 06:24温周瑞
长江大学学报(自科版) 2012年11期
温周瑞
谢 平,徐 军
太湖秀丽白虾繁殖生物学研究
于2004年11月~2005年11月每月采用虾拖网采样对太湖梅梁湾和贡湖湾秀丽白虾(Exopalaemonmodestus)的繁殖生物学进行了研究。结果表明,4月下旬至9月下旬为太湖秀丽白虾的繁殖期,这期间太湖水温变化范围为18.7~32.0℃。6月中旬的繁殖最高峰是由前一年出生的个体产生,而8月的繁殖最高峰则主要是由当年出生的个体所产生。梅梁湾和贡湖湾秀丽白虾性比分别变动在0.76~2.76和0.73~3.47,年平均性比分别为1.38和1.41,卡方检验性比不等于1∶1(Plt;0.01),雌虾多于雄虾。梅梁湾抱卵秀丽白虾生物学最小型体长为26.7mm、体重为0.23g,抱卵量22粒,贡湖湾秀丽白虾生物学最小型体长22.1mm、体重0.13g,抱卵量10粒。梅梁湾、贡湖湾秀丽白虾绝对繁殖力F(卵粒数)与体长、体重之间基本上都有正相关关系。除7月份以外,5月、6月、8月梅梁湾秀丽白虾抱卵虾平均体长、平均体重、绝对繁殖力和相对繁殖力均明显大于贡湖湾秀丽白虾。5月份、8月份梅梁湾秀丽白虾性腺成熟系数分别为(12.2±5.3)%(n=33)、(10.9±4.1)%(n=70),两者没有显著性差异(Pgt;0.05)。发现该结果比1980年太湖秀丽白虾的抱卵量小,大规格虾数量较少,抱卵虾整体偏小,可能与太湖污染加剧及捕捞强度过大有关。
秀丽白虾(Exopalaemonmodestus);繁殖生物学;太湖
秀丽白虾(Exopalaemonmodestus)属十足目长臂虾科白虾属,多生活于大中型湖泊中。在我国淡水水域中,其产量仅次于日本沼虾而居第二位。在大型湖泊中其产量高于日本沼虾,如太湖中秀丽白虾产量可达该湖虾类产量的50%~80%[1-2]。
迄今为止,有关秀丽白虾繁殖生物学的研究报道不多,赖伟等[3]、施炜纲等[4]和胡廷尖等[5]先后报道了太湖秀丽白虾的生物学习性,黄海霞等[6-7]和姜叶琴等[8-10]研究了秀丽白虾生殖系统组织学及染色体核型。此外,谈奇坤等[11]报道过巢湖秀丽白虾繁殖特性。近年来,太湖的水环境发生了很大变化,已从20世纪80年代水质良好的湖泊演变成严重富营养化的湖泊[12]。水环境的持续恶化,加之捕捞强度增大,近年来太湖渔获规格明显小型化[13]。因此,对秀丽白虾繁殖生物学进行研究,对合理保护和利用秀丽白虾资源具有重要意义。
1 材料与方法
1.1 样品采集
分别在太湖的梅梁湾和贡湖湾设立3个采样点,采用GPS定位(图1)。采样时间为2004年11月至2005年11月,每月中旬定期采集1次。用自制的虾拖网(宽3m,网目7mm)采样,每次采样时于每个采样点用虾拖网拖3次,每次5min,捕获的虾全部低温冷藏带回实验室处理。
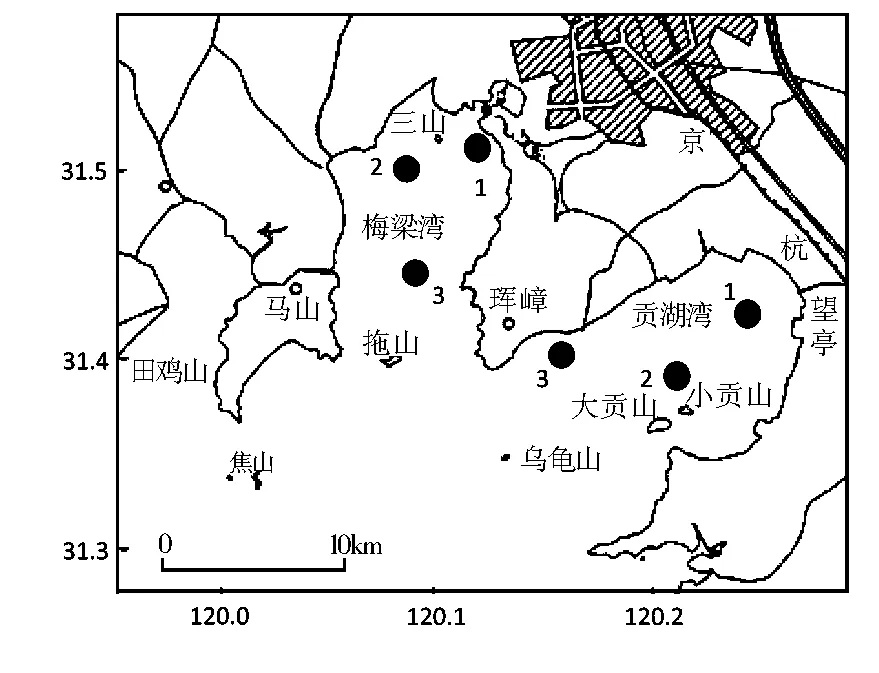
图1 梅梁湾与贡湖湾采样点分布图
1.2 方法
将每次采集到的样品先按谈奇坤[14]的方法鉴定雌雄,确定性比;抱卵虾全部计数,确定雌虾抱卵率。逐一进行体长、体重测定。体长指眼柄基部至尾节末端的距离(精确到0.1mm),体重(湿重)采用吸水纸吸去体表水分后用电子天平称量(精确到0.01g)。共测定梅梁湾秀丽白虾4081只,贡湖湾秀丽白虾3524只。绝对繁殖力和相对繁殖力通过测定抱卵虾的体长和体重,计数其全部抱卵量后计算所得。从样品中出现抱卵虾开始至大部分抱卵虾消失为止的这段时间作为虾的繁殖期。
2005年5月、8月分别将梅梁湾抱卵虾经4%甲醛固定后,取出卵巢和卵粒,用吸水纸吸干水分,称取重量,用性腺重除以去性腺体重,按下式计算性腺成熟系数(gonadosomatic index,GSI)。
GSI=(性腺重/去性腺后体重)×100%。
数据分析采用Statistica 6.0和Excel软件。
2 结果与分析
2.1 性比
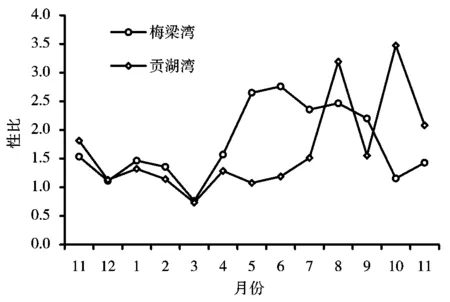
注:月份中11~12为2004年,1~11为2005年(图3同)。
太湖秀丽白虾群体的性比(雌∶雄)在不同月份有着明显变化(图2)。从图2可以看出,梅梁湾与贡湖湾秀丽白虾除3月份雌虾数量少于雄虾外,其他月份均为雌虾数量多于雄虾,梅梁湾和贡湖湾秀丽白虾周年性比分别变动在0.76~2.76和0.73~3.47,年平均性比分别为1.38和1.41,卡方检验表明,雌虾明显多于雄虾(Plt;0.01)。
2.2 繁殖期
经周年采样观察,秀丽白虾3月份至6月份群体平均体长呈增长状态,6月至7月平均体长减小,7月至8月平均体长快速增长,每月雌虾平均体长均大于雄虾平均体长(图3)。4月中旬在梅梁湾和贡湖湾均未发现抱卵虾,但是有部分刚蜕皮的软壳虾,贡湖湾部分雌虾透过头胸甲能看清卵粒,5月中旬采样中有抱卵虾出现。图4显示,5月~9月秀丽白虾群体中雌虾抱卵率的变化情况,梅梁湾秀丽白虾5月抱卵率为30%,6月份上升到82%,7月份为10.5%,8月份升至59%,9月份降至41.7%。贡湖湾秀丽白虾5月抱卵率为72%,6月的抱卵率71.8%,但7月份为25.46%,8月份为45.71%,9月的抱卵率较低,为4%。10月中旬的采样中有个别雌虾抱卵。因此,可确定4月下旬至9月下旬为太湖秀丽白虾的主要繁殖期,这一段时间太湖水温变化范围为18.7~32.0℃。
2.3 繁殖群体的组成
繁殖期间分别测定了梅梁湾和贡湖湾秀丽白虾抱卵虾297只和548只,其平均体长、平均体重的变化见表1。由表1可以看出,从5月~9月,梅梁湾秀丽白虾抱卵虾平均体长、平均体重变化范围为41.4~45.5mm,体重变幅为1.18~1.55g。贡湖湾秀丽白虾抱卵虾平均体长、平均体重分别为30.0~42.3mm、0.44~1.23g。可见,秀丽白虾抱卵虾个体平均体长、体重比雌虾平均体长、体重要大。
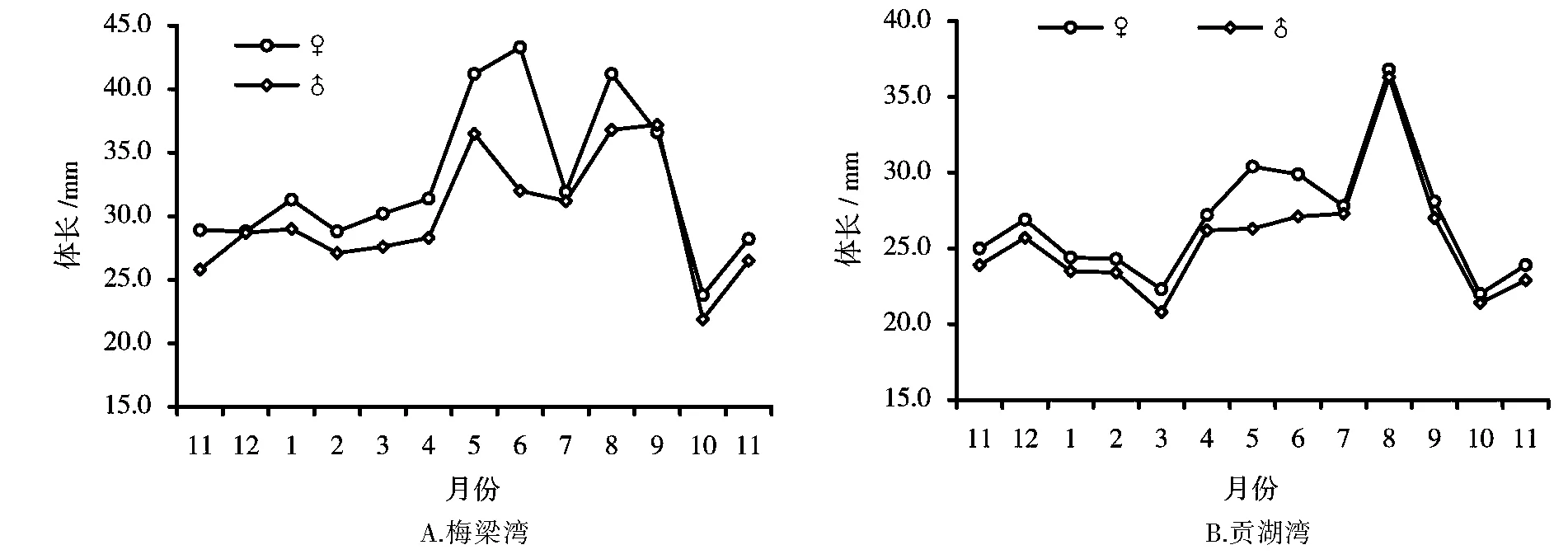
图3 梅梁湾与贡湖湾秀丽白虾平均体长月度变化
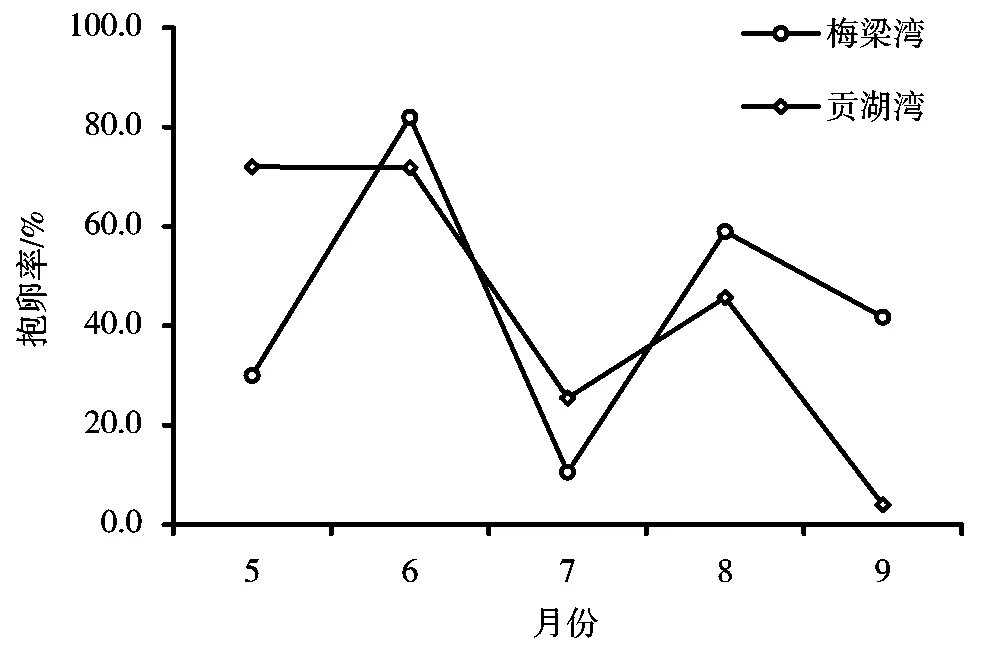
图4 秀丽白虾怀卵率季节变化
将抱卵虾按体长进行分组,组距5mm,计算出每组数量占总数的百分比,作出体长分布图(图5、图6)。
梅梁湾秀丽白虾抱卵群体组成见图5。由图5可见,5月主要是体长45~50mm群体,占60%,其次是40~45mm体长组,占15.56%;6月份主要是体长30~35mm、45~50mm和50~55mm群体,分别占17.95%、34.62%和26.92%;7月主要是体长35~40mm、40~45mm和50~55mm群体,分别占18.75%、43.75%和17.75%;8月份体长40~45mm群体占65.35%,其次是45~50mm群体占22.83%;9月抱卵虾数量减少,组成比例与8月类似。
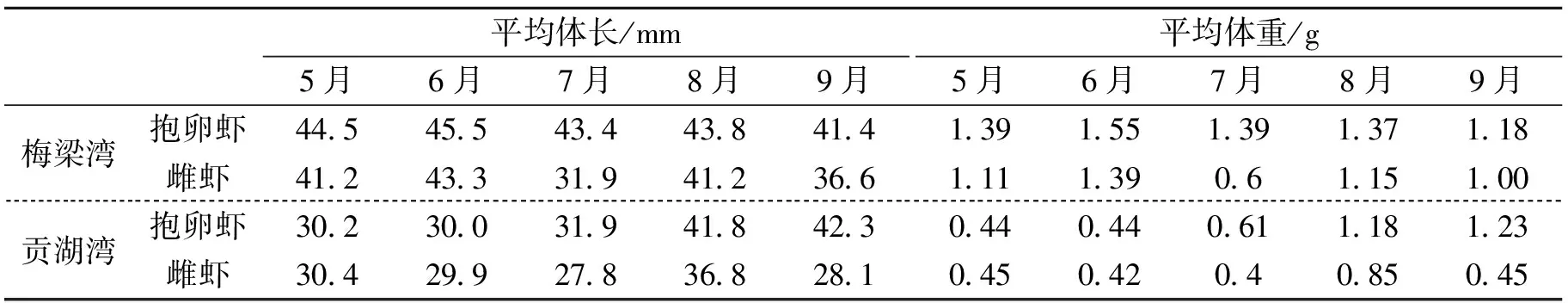
表1 太湖秀丽白虾抱卵虾平均规格
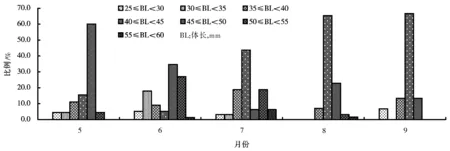
图5 梅梁湾秀丽白虾抱卵虾群体组成
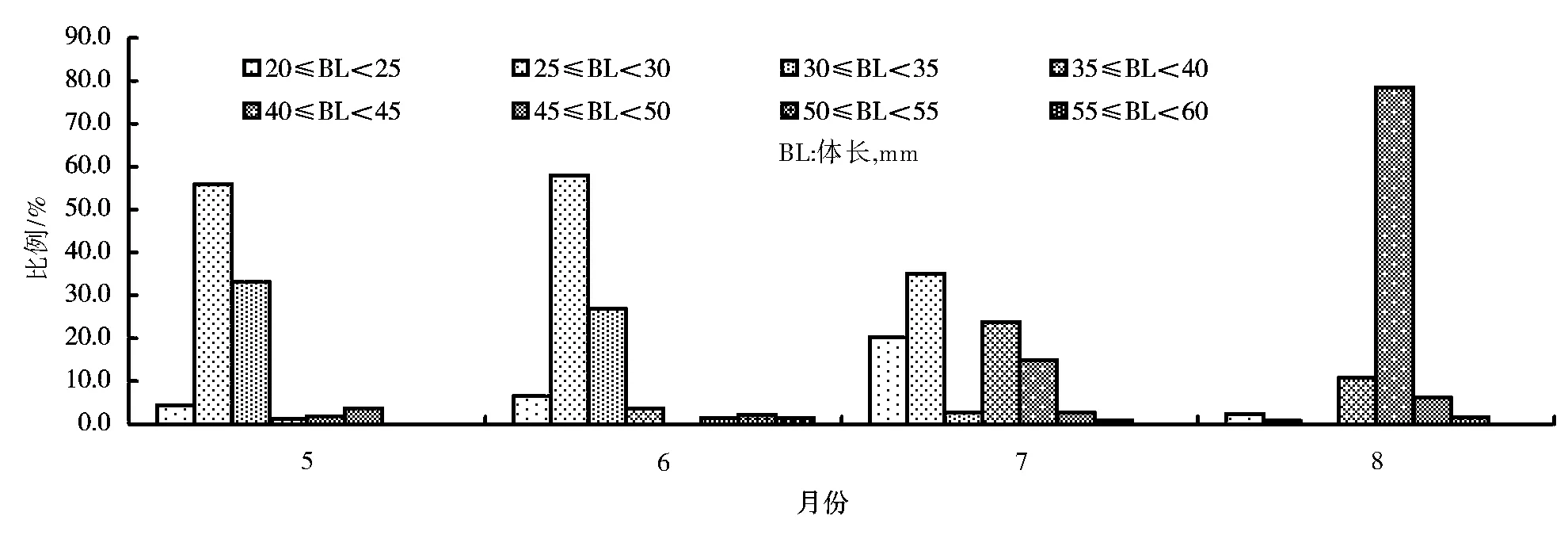
图6 贡湖湾秀丽白虾抱卵虾群体组成
贡湖湾抱卵秀丽白虾群体组成见图6。由图6可见,5月主要是体长25~30mm和30~35mm群体,分别占55.83%和33.13%;6月份抱卵虾组成与5月相似,25~30mm体长组占57.97%,30~35mm体长组占26.81%;7月份抱卵虾体长分布大致集中在两个区间,体长20~30mm占55.27%,体长35~45mm占38.59%;8月份40~45mm体长组占绝对优势,为78.46%,其次是35~40mm体长组,占10.77%;9月份抱卵率较低,抱卵虾较少。
2.4 繁殖力与成熟系数
(1)绝对繁殖力 5月~8月梅梁湾抱卵秀丽白虾的体长范围为26.7~57.3mm(平均44.8±5.5mm)、体重范围为0.23~2.94g(平均(1.45±0.47)g),抱卵量变化于22~364粒之间,绝对繁殖力平均为(153.3±70.0)粒。绝对繁殖力F(卵粒数)与体长L、体重W之间的函数关系为:F=47.481e0.7113W(R2=0.3633);F=10.284e0.0573L(R2=0.3260)。
5月~8月贡湖湾抱卵秀丽白虾的体长范围为22.1~57.2mm(平均(32.4±7.4)mm)、体重范围为0.13~2.96g(平均(0.60±0.50)g),抱卵量变化于10~442粒之间,绝对繁殖力平均(68.5±59.1)粒。绝对繁殖力F(卵粒数)与体长L、体重W之间的相关关系为:F=96.958W+11.006(R2=0.7453);F=3.5926e0.0814L(R2=0.5185)。
(2)相对繁殖力 以单位体重的卵粒数(RW=F/W)和单位体长的卵粒数(RL=F/L)2种方式来表示秀丽白虾的相对繁殖力。
3月~8月梅梁湾RW的变化范围为15.3~245.8粒/g,平均(108.3±43.0)粒/g;RL的变化范围为7.2~71.7粒/cm,平均(33.3±13.9)粒/cm。RL与体长L之间的没有相关性。
3月~8月贡湖湾RW的变化范围为30.4~326.8粒/g,平均(124.3±50.5)粒/g;RL的变化范围为3.7~83.6粒/cm,平均(19.2±11.7)粒/cm。RL与体长L、体重W之间的没有相关性。
5月~8月梅梁湾、贡湖湾抱卵虾繁殖力比较,除7月份以外,5月、6月、8月梅梁湾秀丽白虾抱卵虾平均体长、平均体重、绝对繁殖力和相对繁殖力均明显大于贡湖湾秀丽白虾(表2)。
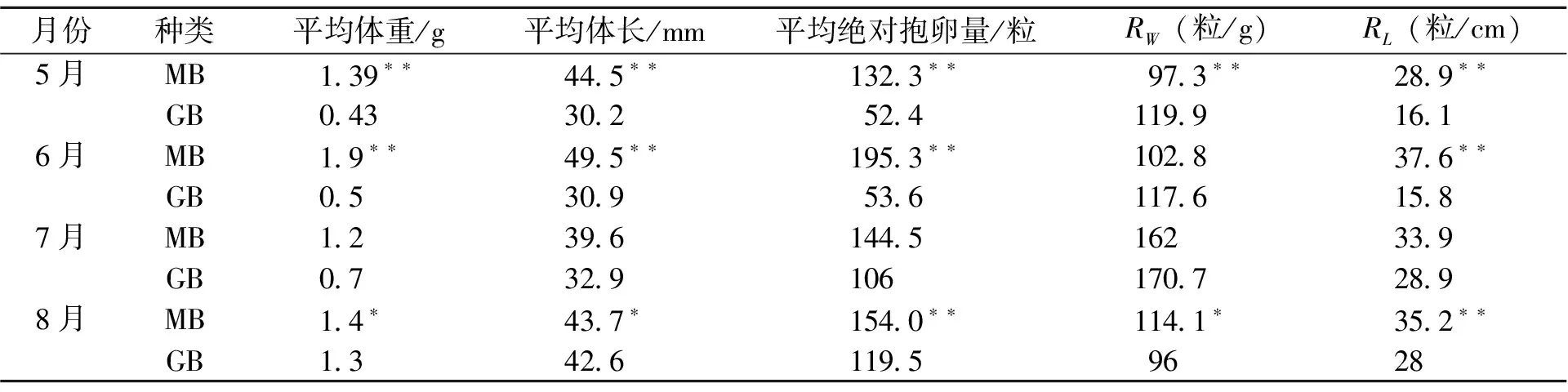
表2 梅梁湾与贡湖湾秀丽白虾繁殖力比较
(3)成熟系数 5月份、8月份梅梁湾秀丽白虾性腺成熟系数分别为(12.2±5.3)%(n=33)、(10.9±4.1)%(n=70),两者没有显著性差异(Pgt;0.05)。
3 讨论与小结
3.1 秀丽白虾的性比
大多数甲壳动物种类因没有性染色体,其性别决定主要受环境影响[15]。如罗氏沼虾(Macrobrachiumrosenbergii)和日本沼虾(M.Janpponese)未发现有异型性染色体[16]。十足类性别受外界环境因子如温度、食物供给、密度等影响较大[17-18]。据报道[15],褐虾属(Crangon)♀性在寒冷季节明显多于♂性,夏季性比约为l∶1,秋季♀性明显少于♂性。拟须虾属(Aristaeomorpha)♀虾4月~5月较多,8月~10月较少。鹰爪虾(Trachypenaeuscurvirostris)春季是♀性多于♂性,而秋季则相反。梅梁湾和贡湖湾秀丽白虾周年性比分别变动在0.76~2.76,0.73~3.47,年平均性比分别为1.38和1.41,卡方检验性比不等于1∶1(Plt;0.01),雌虾多于雄虾。这与赖伟等[3]报道的雌雄性比为1.43相近。太湖秀丽白虾群体产卵持续时间较长(4月~9月),这段时间内水温变化较大(16.8~32.0℃),不同时间孵化的幼体雌雄比例是否与水温有关,值得进一步研究。
3.2 秀丽白虾的繁殖期
经周年采样观察判断,梅梁湾和贡湖湾秀丽白虾繁殖期均为4月下旬至9月下旬。6月份抱卵虾急剧增多,7月份抱卵率均较6月份明显下降,8月份抱卵率显著升高。6月份以前抱卵虾主要是越年虾,7月份越年虾大部分已经抱卵,当年虾个体较小,性成熟个体不多,抱卵虾群体仅由越年虾中未死亡的个体组成,这样就导致雌虾抱卵率有所降低,此后随着当年较早出生的秀丽白虾性成熟并加入繁殖群体,8月份雌虾抱卵率又升高,在6月和8月形成2个繁殖高峰。这与胡廷尖等[5]报道的南太湖地区秀丽白虾繁殖期相似。采样中见到抱卵的同时,淡绿色或棕黄色的卵巢又在继续发育,说明雌虾连续产卵2次以上。
3.3 生物学最小型
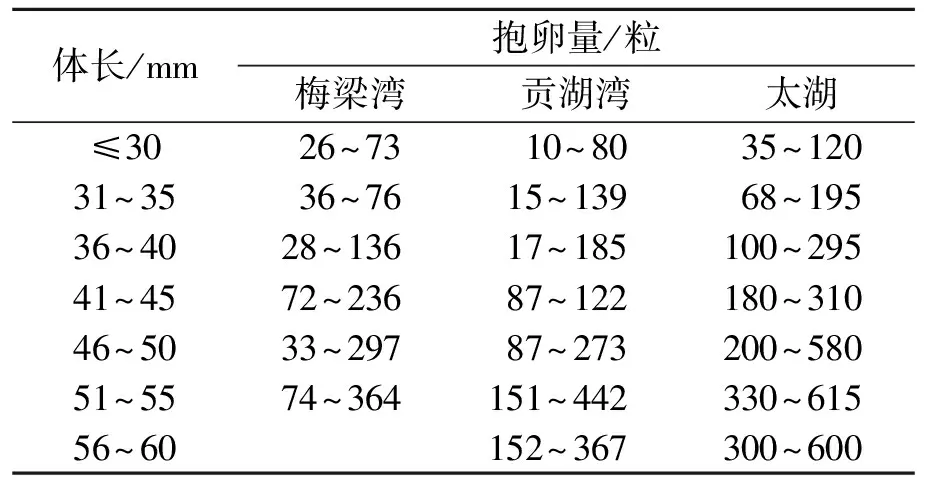
表3 不同体长组秀丽白虾抱卵量比较
梅梁湾抱卵秀丽白虾生物学最小型体长为26.7mm、体重为0.23g,抱卵量22粒,贡湖湾秀丽白虾生物学最小型体长22.1mm、体重0.13g,抱卵量10粒。梅梁湾秀丽白虾生物学最小型大于贡湖湾,可能是由于梅梁湾营养水平比贡湖湾高,饵料生物丰富,秀丽白虾生长速度快,故成熟个体较大。梅梁湾抱卵秀丽白虾生物学最小型与20世纪80年代初的最小体重0.25g相近,但施炜纲等[4]报道1992年8月当年太湖秀丽白虾实验室条件下生物学最小型平均体长27mm,平均体重0.48g,平均怀卵量88粒;同期太湖中的生物学最小型体长31mm,体重1.01g,怀卵量126粒,远大于此次调查的结果,说明目前秀丽白虾有规格趋小的迹象,这与过度捕捞导致的太湖虾类资源小型化的结果相符合。此次采到的最大秀丽白虾抱卵个体梅梁湾为2.94g,贡湖湾为2.96g,仅相当于上世纪80年代初秀丽白虾抱卵虾最大体重5.8g的50%(严生良,粱象秋,金德沂.《太湖虾类的初步研究》(《太湖水产资源调查材料汇编(1980~1981年)》,1981)。
3.4 秀丽白虾的繁殖力
由于梅梁湾抱卵秀丽白虾个体比贡湖湾的抱卵秀丽白虾大,绝对抱卵量也较多,但是梅梁湾RW小于贡湖湾,而RL大于贡湖湾,说明梅梁湾抱卵秀丽白虾肥满度大于贡湖湾。
比较本研究中秀丽白虾与20世纪80年代初太湖秀丽白虾不同体长组抱卵量,发现此次的结果比1980年的抱卵量小,大规格虾数量较少,抱卵虾整体偏小(表3),可能与太湖污染加剧及捕捞强度过大有关。
[1]李新正,刘瑞玉,梁象秋.中国动物志(第四十四卷)[M].北京:科学出版社,2007:61-63.
[2]许志强,葛家春,曹 廷,等.江苏3个秀丽白虾野生群体遗传多样性的AFLP分析[J].江苏农业学报,2009,25(1):160-164.
[3]赖 伟,邓雪怀,陈炳良,等.太湖秀丽白虾种群及生殖习性研究[J].华东师范大学学报(自然科学版),1983,(3):81-87.
[4]施炜纲,严小梅,邴旭文.太湖秀丽白虾生物学与食性的研究[J].湖泊科学,1995,7(1):69-76.
[5]胡廷尖,周志明,黄鲜明,等.秀丽白虾繁殖特性与人工育苗技术的研究[J].中国水产,2001,(1):46-47.
[6]黄海霞,谈奇坤,郭延平.秀丽白虾繁殖生物学研究I.雄性生殖系统的结构与发育[J].安徽师范大学学报(自然科学版),1999,22(3):231-235.
[7]黄海霞,谈奇坤,郭延平.秀丽白虾精子发生的研究[J].动物学杂志,2001,36(2):2-6.
[8]姜叶琴,谢树海,周 琴,等.秀丽白虾染色体核型的初步分析[J].水产科学,2008,27(9):470-472.
[9]姜叶琴,杨万喜.秀丽白虾早期生精细胞结构特征的研究[J].海洋湖沼通报,2003,(2):48-53.
[10]姜叶琴.秀丽白虾卵母细胞不同发育阶段线粒体的变化[J].中国水产科学,2005,12(1):10-l2.
[11]谈奇坤,温耀群,董秀英,等.巢湖虾类资源现状及其利用意见[J].淡水渔业,1985,(3):10-13.
[12]毛新伟,徐 枫,徐 彬,等.太湖水质及富营养化变化趋势分析[J].水资源保护,2009,25(1):48-51.
[13]杨再福,陈立侨,陈 勇,等.太湖渔业资源量变化与对策[J].淡水渔业,2004,34(6):3-5.
[14]谈奇坤.几种淡水虾雌雄鉴别[J].淡水渔业,1991,(5):14-16.
[15]郑 重.甲壳动物的环境型性决定和性比研究[J].台湾海峡,1990,9(3):191-199.
[16]康现江,王所安.高等甲壳动物性别决定机制及其性逆转[J].动物学杂志,1998,33(3):43-46.
[17]Korelainen H.Sex ratios and conditions required for environmental sex determination in animals[J].Biological Review,1990,65:147-184.
[18]Benzie J A H.Penaeid genetics and biotechnology[J].Aquaculture,1998,164: 23-47.
10.3969/j.issn.1673-1409(S).2012.04.007
S917;Q959.233+.63
A
1673-1409(2012)04-S023-05
2012-03-05
863计划项目(2002AA601011);十一五水专项(2008ZX07101-001)。
温周瑞(1966-),男,江西瑞昌人,博士,研究员,研究方向为渔业生态学。
谢 平,E-mailxieping@ihb.ac.cn。
猜你喜欢
天天爱科学(2022年9期)2022-09-15
昆明医科大学学报(2022年1期)2022-02-28
海洋渔业(2021年1期)2021-03-18
水生生物学报(2021年1期)2021-02-04
当代水产(2020年3期)2020-06-15
猪业科学(2018年4期)2018-05-19
猪业科学(2016年1期)2016-03-30
浙江大学学报(农业与生命科学版)(2015年4期)2015-12-15
应用海洋学学报(2014年4期)2014-11-22
中国海洋大学学报(自然科学版)(2014年9期)2014-02-28
