
生育三烯酚生物合成与生理功能研究进展
2012-11-10 06:16范丽萍靳雅欣赵福永长江大学生命科学学院湖北荆州434025
长江大学学报(自科版) 2012年11期
范丽萍,靳雅欣,赵福永 (长江大学生命科学学院,湖北 荆州 434025)
生育三烯酚生物合成与生理功能研究进展
范丽萍,靳雅欣,赵福永 (长江大学生命科学学院,湖北 荆州 434025)
生育三烯酚是天然维生素E的重要组成物质,包含α、β、γ、δ-生育三烯酚4种同系物,仅在植物中有合成。生育三烯酚因具有多不饱和侧链不仅是一种优良的抗氧化剂,而且还具有抗癌、防治心脑血管疾病等重要生理功能。近几年来,对生育三烯酚的生物合成及生理作用进行了广泛地研究,就这些研究成果进行了一个阶段性归纳总结,并就生育三烯酚的开发与利用等问题进行了讨论。
维生素E;生育三烯酚;生物合成;生理功能
维生素E(Vitamin E,VE)是1922年由Evans和Bishop最先发现的能够显著提高老鼠繁殖力的一种“致育因子”,后被命名为“生育酚”。天然VE是一种脂溶性抗氧化剂,包含生育酚(tocopherol)和生育三烯酚(tocotrienol)2大类,每类均由α、β、γ、δ4种同系物组成[1]。天然VE只能由光合自养植物合成,广泛存在于植物的绿色组织和含油种子中[2-4]。VE特有的理化性质使其具有多种生理功能,可增强动物繁殖能力、提高机体免疫能力、防治心脑血管疾病、糖尿病和癌症等,作为抗氧化剂和营养补充剂广泛应用于医药、保健品、食品、化妆品和饲料等行业[5]。同时,植物自身合成的VE在维持其膜系统的稳定性、光合电子传递、胞间信号转导与转录调控等方面也承担着重要角色,高水平表达可显著提高其抗逆能力[6-9]。鉴于天然VE的重要生理功能和广阔的开发应用前景,20世纪中叶就对生育酚的生物合成、代谢及其生理作用机制等方面开展了广泛的研究,尤其是对生物活性最高的α-生育酚研究得最为深入[10]。直到20世纪80年代末,生育三烯酚的降低胆固醇和抗癌特性的发现才逐渐引起研究者的重视。近几年来,生育三烯酚的非抗氧化性生理功能得到了广泛地发掘和研究,为此,笔者就这些研究成果进行了一个阶段性的归纳和总结。
1 生育三烯酚的生物合成与来源
天然VE是通过一系列的酶促反应在叶绿体内膜合成的[11],合成途径受多个关键酶基因(VTE1、VTE2、VTE3、VTE4、VTE5)调控(图1)[12]。生育三烯酚和生育酚的合成均是通过非甲羟戊酸途径和酪氨酸降解途径来获得合成前体物。在单子叶植物中,尿黑酸牻牛儿基牻牛儿基转移酶(HGGT)牻牛儿基牻牛儿基二磷酸(GGDP)和尿黑酸(HGA)为底物合成生育三烯酚前体物2-甲基-6-牻牛儿基牻牛儿基苯醌(MGGBQ),进而进入生育三烯酚合成途径;而在双子叶植物中,尿黑酸植基转移酶(HPT)可以植基二磷酸(PDP)和尿黑酸为底物催化合成生育酚前体物2-甲基-6-植基苯醌(MPBQ)。研究表明,尽管单子叶来源的HGGT和双子叶来源的HPT在氨基酸序列上存在40~50%的相似性,但是它们对底物的亲和力却有显著差别,HGGT与GGDP的亲和力是其与PDP的5倍,而HPT对PDP的亲和力则是其与GGDP的50~80倍[12]。
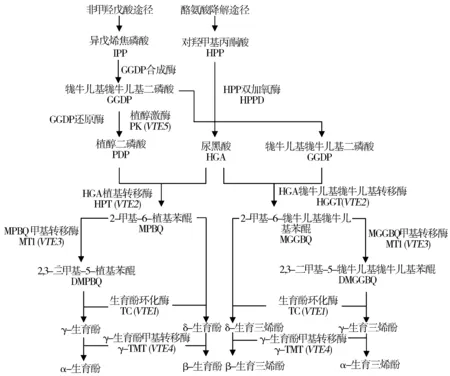
图1 植物维生素E生物合成途径
天然VE广泛地存在于油菜、玉米和大豆等各种油料作物的种子中。研究发现,油菜籽中不论是VE的总含量还是α-生育酚的含量均要高于玉米和大豆,平均含量分别达到了109、42.61mg/100g[3]。核桃仁中的VE含量也比较高,高效液相色谱分析表明,其总含量在8.51~35.28mg/100g之间,α-生育酚含量偏低,仅为0.27~1.53mg/100g[2]。说明未经精炼的植物油是一种人类VE补充的良好膳食来源。
生育三烯酚往往在单子叶植物种子及其植物油中含量较高,如大麦(910mg/kg)、棕榈油(738mg/kg)、米糠(465mg/kg)、燕麦(210mg/kg)、小麦胚(189mg/kg)、椰果(25mg/kg)等,且各种植物中各种生育三烯酚同系物的成份和相对含量也不一致,棕榈油中只含有α、γ、δ-生育三烯酚3种,且γ-生育三烯酚含量最高,达(439mg/kg),而米糠中仅含α、γ-生育三烯酚2种[13]。
2 生育三烯酚的生理功能
Matringe等[9]研究了生育三烯酚在植物中的生理作用。结果发现,生育三烯酚在转HPPD-PDH烟草的幼叶中有积累,并且在低温强光胁迫下可以将光系统Ⅱ的光合抑制和质体膜的过氧化反应维持在一个较低水平,说明生育三烯酚在植物体内可以作为质体膜过氧化反应的有效抗氧化剂。与生育酚不同,生育三烯酚因在侧链上具有3个不饱和双键,有高于生育酚50倍的抗氧化力,对皮肤细胞膜的浸透力也比生育酚要高15倍。4种生育三烯酚同系物中生理活性最高的是δ-生育三烯酚[12]。而生育酚当中,生理活性最高的是α-生育酚,其活性分别是β、γ、δ-生育酚的2倍、10倍、33倍,其生理功能也是最为清楚的[10]。体外条件下,相同浓度的生育三烯酚、α-生育酚和γ-生育酚在质体氧化过程中清除氧自由基的能力相当。但是体内氧自由基的清除主要是α-生育酚,而非γ-生育酚,主要是α-生育酚在组织中的含量是γ-生育酚的10倍左右[14]。人类和高等哺乳动物从食物中摄取的维生素E在小肠内被吸收,然后经过淋巴系统进入体内循环,最后以乳糜微滴形式被转运致肝脏[15]。维生素E是由细胞色素P450进行代谢并最终由尿液排除体外[16]。
2.1 α-生育三烯酚的功能
α-生育三烯酚在色满环R1、R2、R3 3个位置均含有甲基,是在γ-生育三烯酚基础上合成的产物,在大麦中含量最高,达670mg/kg,其最重要的生理功能在于神经保护作用。
中枢神经系统脑组织中富含花生四烯酸等多种多不饱和脂肪酸,花生四烯酸的氧化不稳定性会引起大脑神经细胞的病变。棕榈油中的α-生育三烯酚能够通过酶代谢途径和非酶代谢途径减缓花生四烯酸的氧化进程[17]。谷氨酸盐可激活pp60(c-Src)和12-脂氧合酶(12-lipoxygenase,12-Lox)的表达,是导致神经细胞死亡的重要因素。药理学和遗传学研究表明,12-Lox的酪氨酸在c-Src存在的条件下会快速磷酸化,这种磷酸化对c-Src的抑制因子及α-生育三烯酚非常敏感,纳摩尔浓度的α-生育三烯酚能起到很好的抑制作用。α-生育三烯酚也可直接结合12-Lox及磷酸化的12-Lox而影响其催化活性,说明神经细胞胞质中的c-Src和12-Lox是α-生育三烯酚的作用靶标,α-生育三烯酚可以通过其抗氧化作用和非抗氧化功能起到保护神经的作用[18]。此外,α-生育三烯酚还具有阻止血小板血栓形成和血小板凝集的作用[19]。
2.2 β-生育三烯酚的功能
β-生育三烯酚在色满环R1、R3 2个位置含有甲基,是在δ-生育三烯酚基础上合成的产物,在小麦中含量较高,达33~43mg/kg[20]。目前对其生理功能的认识还很肤浅,仅发现其抑制小牛动脉内皮细胞增殖与微管形成活性高于γ-生育三烯酚和α-生育三烯酚[21]。
2.3 γ-生育三烯酚的功能
γ-生育三烯酚在色满环R2、R3 2个位置含有甲基,是生育三烯酚合成途径中第一个合成的产物,在棕榈油中含量较高,其主要生理功能表现在抗癌和抑制细胞衰老。
徐伟丽等[22]研究发现,γ-生育三烯酚对结肠癌细胞株HT-29具有很强的细胞毒作用,可导致癌细胞中DNA断裂损伤,且与浓度呈一定相关性。孙文广等[23]也证明,γ-生育三烯酚人胃癌SGC-7901细胞生长具有抑制作用,具有剂量和时间效应,推测可能是γ-生育三烯酚导致癌细胞中DNA断裂损伤而诱导的细胞凋亡有关。Makpol等[24]选取了不同年龄(21岁、40岁、68岁)人的成纤维原细胞用H2O2处理产生氧化胁迫,对处理前、后的细胞再用一定浓度(40μmol/L或80μmol/L)的γ-生育三烯酚进行再处理。结果表明,γ-生育三烯酚可以抑制人成纤维原细胞的衰老,主要是通过抗氧化作用显著地阻止端粒长度变短和端粒酶活性降低。相对年幼细胞而言,衰老的人成纤维原细胞中CCND1、RB1、MMP1和IL6基因的表达水平提高。将其与γ-生育三烯酚共培养24h后,衰老的人成纤维原细胞中CCND1、RB1的表达水平会显著降低,且处于G0/G1期的细胞群体减少,而处于G2/M期的细胞群体增加。不论在衰老细胞还是年轻细胞,γ-生育三烯酚均会引起与衰老相关基因表达水平的变化,表明γ-生育三烯酚可以通过调节细胞周期和与衰老相关基因的表达来延缓细胞的衰老[25-26]。
一定剂量的γ-生育三烯酚与其他化学抗癌药物联用可以对乳腺癌细胞的生长与活力起到很好的抑制作用[27-28]。抑制素是3-hydroxy-3-methylglutaryl-coenzyme A(HMGCoA)还原酶的优良抑制剂,是一种良好的抗癌化学药物,但是具有较大毒性。厄洛替尼和吉非替尼也是一种抗癌良药,它可抑制HER/ErbB受体亚型的活性,但是癌细胞中不同EGF受体之间可以形成异源二聚体,只针对其中1个亚型也起不到良好的杀灭作用。低剂量的γ-生育三烯酚与Celecoxib联用可以通过依赖和不依赖COX-2两种机制具有协同抗癌作用,前者主要是抑制了PGE2的表达,而后者主要是减少了ErbB2-4受体表达量[29]。
2.4 δ-生育三烯的功能
δ-生育三烯仅在色满环R3位置含有甲基,其主要生理功能在于抗癌和抑制肿瘤细胞生长作用。
李大鸣等[30]研究发现,一定浓度(gt;30μmol/L)的δ-生育三烯酚对结肠癌SW620细胞的生长具有明显的抑制作用,推测可能是δ-生育三烯酚上调了癌细胞中轴蛋白(Axin)的表达而实现抑制作用的。δ-三烯生育酚还能通过调控Bax、Bcl-2等细胞凋亡相关基因的表达抑制鼻咽癌5-8F细胞的增殖并诱导其凋亡[31]。δ-生育三烯还具有辐射防护作用,其机制可能在于δ-生育三烯可以激活胞外信号传导相关激酶的磷酸化而阻止DNA损伤的产生。δ-生育三烯可以促进rapamycin靶标蛋白的表达以及其下游效应因子4EBP-1的磷酸化,这些均与负责细胞存活与生长的核糖体蛋白S6和翻译调控因子eIF4E转录激活有关[32]。
免疫系统功能的变化导致的多种与年龄相关的疾病的发生与体内一氧化氮的增加密切相关。δ-三烯生育酚可抑制由脂多糖诱导的与衰老和炎症相关基因(IL-1b、IL-1a、IL-6、TNF-a、IL-12、iNOS、VCAM1、ICAM1、COX2、IL-1RA、TRAF1、CD40)的表达,从而抑制一氧化氮的产生[33-34]。生育三烯酚可以通过下调内表皮细胞VEGF受体的表达而减少VEGF诱导的血管的形成,进而抑制肿瘤细胞的生长[35]。生育三烯酚在纳摩尔浓度具有保护神经的作用,在毫摩尔浓度具有抗癌作用。Husain等[36]研究发现,将小鼠100mg/kg剂量口饲δ-生育三烯酚,连续饲喂6周后,胰腺癌组织中的δ-生育三烯酚含量可达(41.8±3.5)nmol/g,可以抑制80%的癌细胞生长。而胰腺中的含量是癌组织中的10倍,没有表现出组织病理变异和毒性。
4 展望
提高植物维生素E含量不仅可以扩大人类和高等哺乳动物天然维生素E的摄取来源,而且还可以提高植物自身的抗逆性,加之维生素E表现出来的重要生理功能,因此如何提高植物中的维生素E含量近来已被植物育种家们提上议事日程。自然界中很多植物本身的维生素E含量就较高,因此可以以这些材料为亲本进行杂交育种,从而选育出高维生素E含量的品种;但是,有些研究[2-4、37-39]表明,维生素E含量的遗传受环境因素影响很大,这给采用常规育种途径培育高维生素E含量的品种带来了一定的困难。然而,目前对于维生素E的生物合成途径已比较清楚,多个关键酶合成基因也已被克隆,因此,采用基因工程手段来提高植物中的维生素E含量也不失为一条理想途径。然而,近十年来采用基因工程手段来提高花生、玉米、油菜等油料作物种子中的维生素E含量的报道屡见不鲜[40-45],但是真正实现商业化生产的品种却没有,可见优良的基因资源仍是制约这一技术的瓶颈。
尽管生育三烯酚体外抗氧化能力是生育酚的50多倍,且具有预防动脉硬化、风湿性关节炎、心脏疾患、脑梗塞、癌症、糖尿病性视网膜症、血管新生病、延缓细胞衰老等重要生理作用,但是人类和高等哺乳动物肝脏中的生育酚结合蛋白(tocopherol-binding protein,TBP)仅对α-生育酚具有高度的专一性,使α-生育酚总是被优先吸收和利用,而生育三烯酚很难被吸收而发挥功效,因此如何促进生育三烯酚的吸收和利用也是一个值得研究的问题。生育三烯酚被认为是开发抗癌疫苗的良好佐剂,与其他抗癌药物联用表现出了良好的抗癌效果[46-48],但是对其分子机制目前还没有深入的认识,广泛开展这方面的研究也是深入利用生育三烯酚的一个重要方面。
[1]Kamal-Eldin A,Appelqvist LA.The chemistry and antioxidant properties of tocopherols and tocotrienols [J].Lipids,1996,31:671-701.
[2]孙 翠.核 桃(JuglansregiaL.)坚果仁维生素 E含量分析及jrMPBQ基因克隆 [D].青岛:山东农业大学.2010.
[3]李禄慧,徐妙云,张 兰,等.不同作物中维生素E含量的测定和比较 [J].中国农学通报,2011,27(26):124-128.
[4]Corzo-Valladaresa P A,Fernández-Martínezb J M,Velascob L.Tocochromanol content and composition inJatrophacurcasseeds [J].Industrial Crops and Products,2012,36:304-307.
[5]Brigelius-Flohe R,Traber M G.Vitamin E:function and metabolism [J].FASEB J,1999,13:1145-1155.
[6]Havaux M,Eymery F,Porfirova S,et al.vitamin E protects against photoinhibition and photooxidative stress inArabidopsisthaliana[J].Plant Cell,2005,17:3451-3469.
[7]Kanwischer M,Porfirova S,Bergmüller E,et al.Alterations in tocopherol cyclase activity in transgenic and mutant plants ofArabidopsisaffect tocopherol content,tocopherol composition,and oxidative stress [J].Plant Physiol,2005,137:713-723.
[8]郭 娟,刘小丽,李晓峰,等.拟南芥VTE1过量表达可以增加维生素E含量和提高烟草植株耐盐性[J].应用与环境生物学报,2006,12(4):468-471.
[9]Matringe M,Ksas B,Rey P,et al.Tocotrienols,the unsaturated forms of vitamin E,can function as antioxidants and lipid protectors in tobacco leaves [J].Plant Physiol,2008,147:764-778..
[10]Engin K N.Alpha-tocopherol:looking beyond an antioxidant [J].Molecular Vision,2009,15:855-860.
[11]Soll J,Douce R,Schultz G.Site of biosynthesis of α-tocopherol in spinach chloroplasts [J].FEBS Lett,1980,112:243-246.
[12]Rebeiz C A,Benning C,Bohnert H J,et al.Advances in Photosynthesis and Respiration(Volume 31):The Chloroplast:Basics and Applications[M].Dordrecht:Springer,2010.
[13]http://www.tocotrienol.org/index.php?option=com_contentamp;view=articleamp;id=49amp;itemid=59.
[14]Wolf G.How an increased intake of alpha-tocopherol can suppress the bioavailability of gamma-tocopherol [J].Nutr Rev,2006,64:295-299.
[15]Fairus S,Nor R M,Cheng H M,et al.Alpha-tocotrienol is the most abundant tocotrienol isomer circulated in plasma and lipoproteins after postprandial tocotrienol-rich vitamin E supplementation [J].Nutrition Journal,2012,11:5.
[16]Brigelius-Flohé R.Vitamin E and drug metabolism [J].Biochem Biophys Res Commun,2003,305:737-740.
[17]Sen C K,Rink C,Khanna S.Palm oil-derived natural vitamin E α-tocotrienol in brain health and disease [J].J Am Coll Nutr,2010,29(3 Suppl):314S-323S.
[18]Khanna S,Parinandi N L,Kotha S R,et al.Nanomolar vitamin E α-tocotrienol inhibits lutamate-induced activation of phospholipase A2 and causes neuroprotection [J].J Neurochem,2010,112:1249-1260.
[19]Qureshi A A,Karpen C W,Qureshi N,et al.Tocotrienols-induced inhibition of platelet thrombus formation and platelet aggregation in stenosed canine coronary arteries [J].Lipids in Health and Disease,2011,10:58.
[20]Panfili G,Fratianni A,Irano M.Normal phase high-performance liquid chromatography method for the determination of tocopherols and tocotrienols in cereals [J].J Agric Food Chem,2003,51:3940-3944.
[21]Miyazawa T,Tsuzuki T,Nakagawa K,et al.Antiangiogenic potency of vitamin E [J].Ann N Y Acad Sci,2004,1031:401-404.
[22]徐伟丽,陈炳卿,王 琪,等.γ-生育三烯酚对人结肠癌HT-29细胞中DNA 损伤作用的研究 [J].疾病控制杂志,2007,11(6):603-606.
[23]孙文广,刘慧坤,乔 羽,等.γ-生育三烯酚对人胃癌SGC-7901细胞的抑制作用及机制 [J].现代肿瘤医学,2012,20(2):230-233.
[24]Makpol S,Abidin A Z,Sairin K,et al.γ-Tocotrienol prevents oxidative stress-induced telomere shortening in human fibroblasts derived from different aged individuals [J].Oxidative Medicine and Cellular Longevity,2010,3:35-43.
[25]Makpol S,Zainuddin A,Chua K H,et al.Gamma-tocotrienol modulation of senescence-associated gene expression prevents cellular aging in human diploid fibroblasts [J].Clinics,2012,67:135-143.
[26]Catalgol B,Ozer N K.Protective effects of vitamin E against hypercholesterolemia-induced age-related diseases [J].Genes Nutr,2012,7:91-98.
[27]Sylvester P W.Synergistic anticancer effects of combined c-tocotrienol with statin or receptor tyrosine kinase inhibitor treatment [J].Genes Nutr,2012,7:63-74.
[28]Kannappan R,Gupta S C,Kim J H,et al.Tocotrienols fight cancer by targeting multiple cell signaling pathways [J].Genes Nutr,2012,7:43-52.
[29]Shirode A B,Sylvester P W.Mechanisms mediating the synergistic anticancer effects of combined γ-tocotrienol and Celecoxib treatment [J].J Bioanal Biomed,2011,3:1-7.
[30]李大鸣,邸金茹,王春花,等.δ-生育三烯酚对人结肠癌SW620细胞抑制作用 [J].中国公共卫生,2010,26(9):1136-1137.
[31]曾 攀.d-δ-三烯生育酚对鼻咽癌细胞增殖与凋亡的影响及分子机制 [D].桂林:桂林医学院,2011.
[32]Li X H,Fu D,Latif N H,et al.δ-tocotrienol protects mouse and human hematopoietic progenitors from γ-irradiation through extracellular signal-regulated kinase/mammalian target of rapamycin signaling [J].Haematologica,2010,95:1996-2004.
[33]Qureshi A A,Tan X Y,Reis J C,et al.Suppression of nitric oxide induction and pro-inflammatory cytokines by novel proteasome inhibitors in various experimental models [J].Lipids in Health and Disease,2011,10:177.
[34]Qureshi A A,Tan X Y,Reis J C,et al.Inhibition of nitric oxide in LPS-stimulated macrophages of young and senescent mice by δ-tocotrienol and quercetin [J].Lipids in Health and Disease,2011,10:239.
[35]Wells S R,Jennings M H,Rome C,et al.Alpha-,gamma and delta-tocopherols reduce inflammatory angiogenesis in human microvascular endothelial cells [J].J Nutr Biochem,2010,21:589-597.
[36]Husain K,Francois R A,Hutchinson S Z,et al.Vitamin E δ-tocotrienol levels in tumor and pancreatic tissue of mice after oral administration [J].Pharmacology,2009,83:157-163.
[37]Li H Y,Liu H C,Han Y P,et al.Identification of QTL underlying vitamin E contents in soybean seed among multiple environments [J].Theor Appl Genet,2010,120:1405-1413.
[38]Marwede V,Schierholt A,Mollers C,et al.Genotype×environment interactions and heritability of tocopherol contents in canola [J].Crop Sci,2004,44:728-731.
[39]Almeida J,Quadrana L,Asís R,et al.Genetic dissection of vitamin E biosynthesis in tomato [J].J Experimental Botany,2011,62:3781-3798.
[40]黄智明,翁海波,席 宇,等.转入HPT1基因的油菜种子中维生素E 含量的提高 [J].植物生理学通讯,2006,42(5):888-890.
[41]刘风珍,万勇善,王洪刚.γ-维生素E甲基转移酶基因转化花生研究 [J].中国粮油学报,2005,20 (1):61-64,68.
[42]Cahoon E B,Hall S E,Ripp K G,et al.Metabolic redesign of vitamin E biosynthesis in plants for tocotrienol production and increased antioxidant content [J].Nature Biotechnology,2003,21:1082-1087.
[43]Falk J,Andersen G,Kernebeck B,et al.Constitutive over-expression of barley 4-hydroxyphenylpyruvate dioxygenase in tobacco results in elevation of vitamin E content in seeds but not in leaves [J].FEBS Lett,2003,540:35 -40.
[44]Van Eenennaam A L,Li G,Venkatramesh M,et al.Elevation of seed [alpha]-tocopherol levels using plant-based transcription factors targeted to an endogenous locus [J].Metab Eng,2004,6:101-108.
[45]Kumar R,Raclaru M,Schü Beler,et al.Characterisation of plant tocopherol cyclases and their overexpression in transgenicBrassicanapusseeds [J].FEBS Lett,2005,579:1357-1364.
[46]Blum S,Vardi M,Brown J B,et al.Vitamin E reduces cardiovascular disease in individuals with diabetes mellitus and the haptoglobin 2-2 genotype [J].Pharmacogenomics.2010; 11:675-684.
[47]Abdul Hafid S R,Radhakrishnan A K,Nesaretnam K.Tocotrienols are good adjuvants for developing cancer vaccines [J].BMC Cancer,2010,10:5.
[48]Yang Z H,Lee M J,Zhao Y,et al.Metabolism of tocotrienols in animals and synergistic inhibitory actions of tocotrienols with atorvastatin in cancer cells [J].Genes Nutr,2012,7:11-18.
10.3969/j.issn.1673-1409(S).2012.04.011
Q566
A
1673-1409(2012)04-S041-05
2012-03-20
湖北省教育厅资助项目(Q20101302)。
范丽萍(1988-),女,海南文昌人,现从事生物技术研究。
赵福永,E-mailfyzhao@yangtzeu.edu.cn。
猜你喜欢
昆明医科大学学报(2020年12期)2021-01-26
抗癌之窗(2020年1期)2020-05-21
特别健康(2018年9期)2018-09-26
猪业科学(2018年4期)2018-05-19
红土地(2018年12期)2018-04-29
广东饲料(2016年4期)2016-12-01
中国卫生(2016年12期)2016-11-23
中国现当代社会文化访谈录(2016年0期)2016-09-26
国外医药(抗生素分册)(2016年3期)2016-07-12
