
浙江省海洋外来生物入侵影响与控制策略研究*
2012-11-06 09:32陈全震曾江宁
海洋开发与管理 2012年5期
高 瑜,陈全震,曾江宁
(国家海洋局第二海洋研究所 杭州 310012)
浙江省海洋外来生物入侵影响与控制策略研究*
高 瑜,陈全震,曾江宁
(国家海洋局第二海洋研究所 杭州 310012)
浙江省是我国遭受海洋生物入侵较为严重的地区之一。文章探讨了浙江海洋生物的入侵途径、外来物种成功入侵的机制及入侵种对生态系统的影响,同时给出了控制海洋生物入侵的策略,以期能更好地保护海洋生物多样性,减少外来生物入侵灾害的发生。
外来海洋生物;入侵机制;生态系统;控制与管理
1 浙江省海洋外来物种入侵名录
浙江省海域辽阔,海洋资源丰富。但是由于近年来浙江海洋运输带来的外来生物以及对海洋动物、植物的不慎引进,导致了浙江海洋环境面临严峻的入侵压力。根据2004—2009年的文献资料,总结出浙江沿海的生物入侵种统计如表1所示。

表1 浙江省海洋外来物种入侵统计表

续表
2 浙江省外来海洋生物入侵途径
入侵生物在无人类介入的情况下,生物在生物区间、大陆间和岛屿间远距离传播也可能发生,但这种入侵只是小概率事件[8]。浙江海洋外来动植物入侵途径可大致分为有意引入导致的生物入侵和无意引入导致的生物入侵。
2.1 有意引入
有意引入的海洋动植物通常是由于海水养殖的需要而引入的经济价值高或性状优良的物种[9],或者是为改善被破坏的海洋生态环境而引进的滩涂植物等。
2.1.1 潮间带植物引种
为抵御风浪、保护海岸,中国自20世纪60年代以来,先后引进了大米草、互花米草、狐米草和大绳草4个种[10-11],这些外来物种对海洋湿地生态系统有直接的影响,也与滩涂养殖业的滩涂开发利用紧密相关[1]。
2.1.2 养殖品种的引进
人们出于经济或其他目的,从国外引入具有经济价值或观赏价值的物种,以提高当地养殖或种植的品种多样性,提高经济效益。例如,眼斑拟石首鱼(美国红鱼)[5],由于其生长速度快、适应能力强和食性广泛等特点,我国于1991年从美国德克萨斯州引进并迅速推广,虽产生了一定的经济效益,但由于管理不善,导致其逃逸事故不断发生。目前浙江省大量养殖的美国红鱼,由于喜食其他鱼类,已对象山港的生物结构造成了一定程度的破坏[12-13];此外,我国为振兴对虾养殖业,引入了国外的养殖品种凡纳滨对虾,但伴随其引入的还有桃拉病毒[14]。2007年夏天,舟山、嘉兴、宁波、杭州和绍兴5个地区,桃拉病毒发病面积超过2 000 hm2,造成了巨大的经济损失。
2.2 无意引入
这种引入方式通常是伴随着进出口贸易、海轮或入境旅游在无意间被引入的[15]。
2.2.1 船底携带外来附着生物
自航海时代以来,所有的远洋船舶底部都附着大量的污损生物,这些污损生物随着船舶航行扩展了生存空间,成为新栖息地的外来物种。如,我国海洋入侵种沙筛贝,估计就是通过巴拿马运河进入太平洋和印度洋,由船只带入我国的台湾和香港地区[16],再传入我国东南海域,形成优势种,对当地海洋生态系统造成不利的影响。
2.2.2 压舱水携带外来生物
在过去的几十年里,为了使船体行驶平稳,人们常常在船底堆积上石子、沙砾和铁等重物,现在则直接抽入海水。货船出发时,人们将海水作为压舱水抽上来,抵达目的地后再将其就地排放,这样压舱水中成活的生物体或孢子等休眠体就在新栖息地繁殖。根据《宁波港入境船舶压舱水携带浮游生物的调查与分析》,宁波口岸随机抽取的52艘国际航行船舶进行压舱水浮游生物的调查结果显示:85%以上的压舱水样中可检出至少1种浮游水生生物种,其中最多的一份压舱水样中共检出16种浮游水生生物,并检出4种外洋种与14种与赤潮发生密切相关种。由此可见,压舱水携带外来生物的概率非常高,而浙江省目前关于此方面的研究工作还相对薄弱[4]。
3 海洋生物入侵机制
虽然由于人类运输和贸易的不断发展而导致了大量物种进入新的环境中,但有很多种在进入到新环境的途中就已死亡,或者在到达新环境之后漫长的适应期内灭绝,只有少数种类可以在新环境中生存并且繁衍后代。研究成功入侵种的生物入侵机制有着重要的理论和实践意义。从理论上说,了解入侵种的侵入机制我们可以知道种群的生活进化史以及生物群落是如何集聚的;从实践上说,它可以提供给我们最为有效地防止生物入侵的方法及手段[17]。
3.1 海洋生物入侵过程
Williamson把生物入侵的各个环节划分为3次转移,分别为逃逸、建群以及入侵(图1),他认为每次转移成功的概率在5%~20%之间,大约是10%,称为“十分之一法则”[18]。
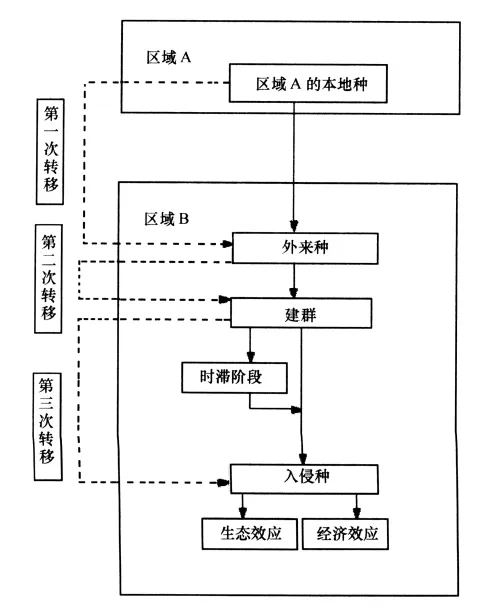
图1 生物入侵链式过程
外来生物入侵发生发展过程类似于Logistic增长模型,大部分外来种在成为入侵种之前都会经历一段或长或短的生长停滞期,而在此阶段,由于其种群数量少且分布分散,常规手段对其无法检出[19]。当外来种成功渡过生长停滞期后,其生长繁殖速度非常迅速,以致在人们发现时已经成为入侵种[20]。
入侵种的扩展主要依靠两种途径:一是通过繁殖使种群向周围空间扩散;二是借助载体传播,可以是远距离的、跳跃式的,主要是通过动物尤其是人的活动携带帮助外来物种扩展[21]。有学者试图通过模型模拟入侵种传播的一般过程[22-23],但其结论还有待证实。
3.2 海洋生物入侵成功机制
在所有外来物种中,既然只有很小一部分物种成为入侵种,这就涉及入侵生物学中的核心问题——哪些物种更易形成入侵种?生物学家Baker针对入侵的杂草性植物物种,总结了12条生活史特征,可以概括为:具有较强的生殖能力;从种子发育到性成熟的周期短;对不良环境的耐受性强等[24]。
空白生态位(vacant niches)假说在一定程度上说明了生物入侵的缘由。许多物种到了一个新环境中,如果当地物种多样性贫乏,没有被占领的、可为入侵种提供的空生态位较多,那么外来种就更易度过生长停滞期而发生爆炸性增殖。但是,在现实中证明空缺生态位的存在极为困难[17]。
群落丰富度假说(community species richness),自1958年Elton首次提出以来成为入侵生态的中心理论之一,他认为群落对外来种入侵的抵抗性与群落中物种数量(即物种丰度)成正比[25]。该假设实际上是由空缺生态位假设推论而来,也就是说,物种丰富的群落为新来者提供空缺生态位的概率不多。
天敌缺乏也是生物入侵成功的重要原因。许多入侵种以种子、孢子、卵或其他休眠形式被带到新地点后,其在本地的竞争者、捕食者和寄生者等并没有随之到达[17],因此这种“天敌缺乏”的状况使得外来种生存空间更大,使得建种群的数量大幅增加,最终暴发成灾[26]。
群落生态学假说则是至今考虑得最为全面的一个假说[27]。该假说是在生态位这一概念的基础上提出的,它把“生态位机遇”作为阐述群落可侵入性的着手点。该假说认为,3种因素影响着外来种的生长速率:资源、天敌和物理环境,他们均有随时空变化的特性,物种对这3种因素及其时空变化的反应决定着其入侵能力。该假说将天敌逃避机遇和资源利用机遇结合起来分析,提出成功的入侵者正是利用各种机遇的胜者,它在群落中缺乏专性甚至兼性天敌,波动的资源正好被它利用,本地竞争者对资源的利用不如它,且兼性天敌对其控制力很强,使其无法与入侵者竞争,物理环境也对其有利。显然,这一假说结合了以上多个假说的观点,最近的生态位理论也阐明,高的物种多样性导致低的生态位机遇(入侵抵抗性)。可以预测,以上各种假设、假说尤其是群落生态学假说将成为入侵生态学(invasion ecology)的主要内容。
4 海洋入侵种对生态系统的影响
4.1 入侵种对物种多样性的影响
外来物种可通过竞争或占据本地物种生态位来排挤本地物种,进而形成单优势群落,导致生态系统结构失衡、生态功能退化。生态系统服务功能的衰退具有持久性和不可逆转性,其生态后果与损失难以用经济数字来估计[28]。如,浙江滩涂的入侵植物互花米草及大米草已对当地生物多样性产生了严重影响。宁波市宁海县双盘涂和蛇蟠涂两地总面积约49.13 km2,最近几年几乎全部被茂盛的大米草侵占,土著种面临严峻的生态危机;1989年,温州市首先在龙湾区引进互花米草,根据2007年卫星遥感影像,温州沿海互花米草分布面积达13.16 km2,比2003年增加了6.29 km2,滩涂上的互花米草为绝对优势种,基本上为单一群落[15]。
生态学家警告,由于地球上人类活动的加剧导致的大规模生物入侵,将会创造一个“超大陆”,必将导致生物多样性灾难性的丧失,全球生物多样性呈现均质化,地区间差异缩小,大多数生物群落将由少数几种入侵能力强的物种所占据[29]。
4.2 入侵种对遗传多样性的影响
有些引入种与当地动植物区系中的某些物种有紧密的亲缘关系,当外来种同当地种发生杂交时,独特的基因型可能从当地种群中消失,物种分类的界限变得模糊不清[8],另外,入侵种与土著种杂交还可能导致种群后代适应性下降、有害种由于杂种优势获得优良性状造成生态危害以及产生杂交群体和广泛的基因渗透,通过“基因污染”而导致本地种的灭绝[30]。
4.3 入侵生物引发赤潮或带入病原微生物
我国沿岸海域的入侵赤潮生物绝大部分属于广布性种类,对海洋生态系统环境的适应性强、分布广[31],一旦环境适宜,就会暴发赤潮。东海是我国赤潮暴发的高频海域,据2009年《中国海洋灾害公报》显示,东海发生赤潮灾害的次数和面积分别占全海域的63.2%和46.5%[32],赤潮的频繁发生,会给我国海洋生态环境以及渔业生产带来极大危害。如,入侵种链状亚历山大藻为能产生麻痹性贝毒(PSP)的种类,微小原甲藻为能产生腹泻性贝毒(DSP)的种类。PSP毒性很强,100 g贝肉中,约500 mg的麻痹性贝毒就可致人死亡[33],另有实验表明,在30 d的实验中赤潮浓度的微小原甲藻对幼贝的致死率在14 d内即达到100%,30%赤潮浓度的原甲藻对幼贝的致死率在22 d内也达到了43%[34]。
外来物种在引入的过程中极可能携带病原微生物,而由于当地动植物对其几乎没有抗性,因此很容易引起病害流行,甚至可能对人类造成严重的伤害[9]。如,笔者提到的凡纳滨对虾携带的桃拉病毒,另外,红鳍东方鲀常携带吻唇溃烂病毒,当水温高于25℃时,则可能出现病发高峰。然而,由于我国对海洋微生物入侵的研究仍处于起步阶段,对海洋微生物的入侵危害掌握还很有限[35],再加上我国目前的检验检疫措施又难以及时发现和阻隔,一旦有病原微生物侵入,带来的则是巨大的生态风险。
5 浙江省海洋外来物种入侵控制策略
5.1 入侵生物治理方案
外来物种入侵以后,对生态系统造成严重的破坏,使生态系统的结构与功能发生改变,抵抗性、自我更新能力和恢复能力弱化。而目前对外来入侵种的治理,尚未有成熟的治理技术,但总结成功的案例,主要有如下几种方式。
5.1.1 人工防治
对于那些刚刚传入、定居,但还没有大面积扩散的入侵生物,人工防治可以在短时间内迅速清除,但对于那些已经大面积扩散的入侵种,机械手段则需要大量的人力、物力,投入大、见效慢,并不是最为经济的方式。
5.1.2 综合防治
将生物、化学、机械和人工等单项技术融合起来,发挥各自优势,弥补各自不足,达到综合控制入侵生物的目的。有害生物综合治理(IPM)是当前国际上普遍认可并广泛采用的有害生物治理策略,该技术从受损生态系统的总体出发,根据有害入侵生物与环境之间的关系,发挥自然因素的作用,因地制宜地协调各种措施,将有害生物控制在经济损害水平之下[36]。如,治理典型入侵生物大米草,只利用单一的方法很难将其彻底铲除。在大米草的入侵初期,可采用人工、机械以及化学的方法进行防治;为维持长期效果,则可运用有效的生态学治理技术,利用天敌进行生物防治,即选用竞争力强的本地物种与大米草竞争,加速大米草的自然演替,寻求新的生态平衡[37]。
5.1.3 资源化利用
有些外来种虽然给当地生态环境带来了恶劣的生态负效应,但本身却有一定的经济价值,若开发得力,则能变废为宝。如,侵蚀海岸滩涂的互花米草属于C4植物,能有效利用二氧化碳和光能,且在整个生长季,其平均净光合速率达24.44μmol CO2m-2/s,是土著滩涂植物芦苇的1.54倍[38]。正因如此,互花米草也具有很高的热值,据钦佩等的研究表明,在苏北地区互花米草平均热值达16 691 J/g。其贮能在9月达最高,为99.52×103kJ/m2,相当于3.39 kg标准煤[39]。目前,互花米草在沿海农村地区作为生活燃料,但最近有研究表明,互花米草直接燃烧效率不高。如果能用其进行沼气生产,则能达到比较满意的利用率[40]。利用互花米草生物质能资源可以作为控制其恶性扩张的较为有效和实用的途径,具有经济和生态双重效益[41]。
另外,如入侵生物红鳍东方鲀中富含大量胶原蛋白,可以提纯、精炼后加入护肤品中,可以具有美容及延缓衰老等功效。
5.2 入侵生物管理方案
生物入侵不仅仅是一个科学问题,也是一个管理问题。在美国,农业科技委员会和农业部领导着“生物入侵”问题的管理与研究;新西兰、澳大利亚等国也相继颁布了相关政策法令,加强了对外来入侵生物的管理。而我国对入侵生物的管理起步晚,目前,针对国内对生物入侵管理的缺失提出以下建议。
5.2.1 健全完善相关法律法规
1996年,美国政府就颁布了《国家入侵物种法》,统一管理入侵物种问题,这部法令是迄今为止国际上专门针对外来种的最为全面的国家法令。而目前,我国无论是国家层面,还是省级层面,还没有针对外来入侵物种的专项法规与条例。各级政府和部门的相关法规也并不完善,有关外来入侵物种管理的条文多分散于不同的法律当中,造成外来物种的管理相对混乱和难以统一。对浙江省而言,有必要建立鱼类引种和野外放生的登记制度,为以后的管理和研究提供翔实可靠的资料。同时参照国外比较成熟的法律系统和经验等,以立法手段设立相应的奖惩措施,私自引种的行为要为其所造成的国家和生态环境的损失付出代价。
5.2.2 建立管理体系,加强管理职能
成立由海洋局和环保部门牵头、多部门协作的专门机构,负责组织制定有关防止海洋有害外来物种入侵的方针、政策、方法、规划和应急方案,开展外来有害物种普查及有关风险进行评估工作等。同时,加强基层监管体系建立,可依托现有渔政、水产技术推广体系建立国家级、省级和县级3级海洋有害外来物种入侵的监控体系,进行时时监控,形成海洋有害外来物种入侵的跟踪预警体系,并对危害最严重的海洋入侵物种进行专项治理[42]。
5.2.3 控制引种规模
如果引种是必要的,在引种之前必须进行生态风险评估,另外还应注意引进种群的大小。因为小种群会出现较大的遗传漂变和近亲繁殖,使不同基因库的群体差异性降低。这样,种群对新环境无法适应而导致种群规模萎缩,在种群规模萎缩的同时,这些遗传问题又会进一步加剧,形成灭绝漩涡。这样小种群因遗传退化和无法进行适应性调整则更易趋于灭绝[9],这可作为防止外来物种成为入侵种的有效手段。
5.2.4 加强国际合作
当前,我国外来海洋生物入侵的数据、资料比较零散,也缺少有效、准确、客观和清晰的评价,这对我国外来海洋生物入侵的立法、监督以及防治无疑是一个巨大的阻碍。因此,应加强国际合作,在广泛的国际交流基础上,制订对共同问题具有共同兴趣的国家合作计划,通过不同渠道,争取国际机构的支持,申请多边与双边的合作课题,在入侵物种风险预警与管理、传统生物防治技术、生态修复技术、区域性综合治理技术以及控制策略与技术等方面开展全面合作[43]。
5.2.5 重视公众教育
通过普及海洋环保知识,提高公众的海洋生态安全意识,杜绝没有经过入侵风险分析的私自引种,减少盲目的放生行为,可在很大程度上达到预防的效果。
6 结束语
海岛作为海洋的一部分,其生物入侵的防控也刻不容缓。2011年2月25日,经国务院批复,浙江海洋经济发展示范区规划上升为国家战略。同年4月13日,国家海洋局公布了我国第一批开发利用无居民海岛名录,其中浙江31个。所以新一轮海洋开发势必增加了浙江海岸带与海岛外来生物入侵的风险概率,有必要未雨绸缪,在优化海洋经济发展布局、打造现代海洋产业体系、构建“三位一体”港航物流服务体系、完善沿海基础设施网络的同时,加强海洋生态文明建设。在“十二五”规划之初,将外来生物管理、入侵灾害预防和海岛生物入侵风险管理等内容纳入涉海类专项规划,并进一步构建系统的法律法规,培养外来海洋生物监测和管理的科技人才,加强外来海洋生物入侵科普教育工作。从法律、生态等科学层面和生物控制、理化防范等技术层面推进浙江省海洋生态文明建设。
应该清楚地认识到,不论是陆地还是海洋,外来入侵的生物正在不断增加,不断地对目前脆弱的生态环境构成直接或者潜在的侵蚀。由于生态系统的复杂性,外来入侵生物带来的危害将会通过食物链、物质循环和能量流动等环节放大,从而影响到整个生态系统。一旦生态系统的结构和功能发生改变,将很难进行修复。这就决定了生物入侵的过程是短暂的,而对生态系统和生物多样性的修复工作却是长期的。入侵物种造成的危害是显而易见的,但其潜在的危害性是较难预见的。对于浙江省海洋生物入侵,要广泛调查,收集数据,建立海洋生物入侵数据库和预警系统。它是最紧迫的基础工作之一。因为利用国内外的数据库,探索生物入侵的规律,明确对浙江海域最具威胁性的生物物种,它们在浙江最有可能生存和暴发的区域,将实实在在地、大幅度地提高浙江对海洋生物入侵的预警那能力。这是很重要的,因为对于生物入侵,预防比治理更为关键。
[1] 徐海根,王建民,强胜.外来物种入侵·生物安全·遗传资源[M].北京:科学出版社,2004:52-77.
[2] 徐海根,强胜.中国外来入侵物种编目[M].北京:中国环境科学出版社,2004:46-48.
[3] 郑剑宁,裘炯良,尤传明,等.国际航船压舱水中浮游水生物携带情况调查[J].中国公共卫生,2006,22(7):874-875.
[4] 郑剑宁,裘炯良,薛新春.宁波港入境船舶压舱水中携带浮游生物的调查与分析[J].中国国境卫生检疫杂志,2006,29(6):358-360.
[5] 李家乐,董志国,李应森,等.中国外来水生动植物[M].上海:上海科学技术出版社,2007:153.
[6] 赵淑江,朱爱意,张晓举.我国的海洋外来物种及其管理[J].海洋开发与管理,2005,22(3):58-66.
[7] 刘红卫,林志凌,苏华轲,等.广东省外来物种入侵现状及其生态环境影响调查[J].生态环境,2004,13(2):194-196.
[8] MOONY H A,CLELAND E E.The evolutionary impacts of invasive species[J].Pro Acad USA,2001,98(10):5446-5451.
[9] 杨圣云,吴荔生,陈明茹,等.海洋动植物引种与海洋生态保护[J].台湾海峡,2001,20(2):259-265.
[10] 王蔚,张凯,汝少国,等.米草生物入侵现状及防治技术研究进展[J].海洋科学,2003,27(7):38-42.
[11] 陈中义,李博,陈家宽,等.米草属植物入侵的生态后果及管理对策[J].生物多样性,2004,12(2):280-289.
[12] 薛利建.网箱养殖美国红鱼逃逸对浙江海域自然生态影响的初步研究[D].青岛:中国海洋大学,2005.
[13] 金海卫,徐汉祥,许文军,等.浙江自然海区逃逸美国红鱼食性研究[J].海洋水产研究,2008,29(1):103-108.
[14] 赵建东.外来海洋生物入侵令人担忧[N].中国海洋报,2009-05-22(A1).
[15] 浙江省环境保护厅.浙江省外来入侵物种调查报告[R].2010.
[16] 黄宗国,蔡如星.海洋污损生物及其防除[M].北京:海洋出版社,1984:242-250.
[17] MACK R N,SIMBERLOFF C D,et al.Biotic invasions:causes,epidemiology,global consequences and control[J].Ecological Applications,2000,10(3):1-20.
[18] WILLIAMSON M.Biological invasions[M].London:Chapman &Hall,1996,244.
[19] 徐承远,张文钜,卢宝荣,等.生物入侵机制研究进展[J].生物多样性,2001,9(4):430-438.
[20] CAREY J R.The incipient mediterranean fruit fly population in California:implications for invasion biology[J].Ecology,1996,77(6):1690-1697.
[21] COHEN A N,CARLTON J T.Accelerating invasion rate in a highly invaded estuary[J].Science Magazine,1998,279(5350):555-558.
[22] SHAROV A A,LIEBHOLD A A.Model of slowing the spread of gypsy moth(Lepidoptera:Lymantriidae)with a barrier zone[J].Ecological Applications,1998,8(4):1170-1179.
[23] SUAREZ A V,HOLWAY D A,CASE T J.Patterns of spread in biological invasions dominated by long-distance jump dispersal:Insights from Argentine ants[J].Proceedings of the National A-cademy of science USA,2001,98:1095-1100.
[24] BAKER H G.The evolution of weeds[J].Annual Reviel of Ecology and Systematics,1974(5):1-24.
[25] ELTON C S.The ecology of invasions by animals and plants[M].London:Metheun,1958:181.
[26] 达良俊,田志慧,王晨曦,等.从生态学角度对生物入侵的思考[J].自然杂志,2007,29(3):152-167.
[27] SHEA K,CHESSON P.Community ecology theory as a framework for biological invasions[J].Trends in Ecology and Evolution,2002,17(4):170-176.
[28] 万方浩,谢丙炎,褚栋.入侵生物学:管理篇[M].北京:科学出版社,2008:149-223.
[29] MOSSELER A.Red pine:a model for the loss of genetic diversity in trees[M].Netherland:SPB Academic Publishing,1995:359-370.
[30] 向言词,彭少麟.生物入侵及其影响[J].生态科学,2001,20(4):68-72.
[31] 赵冬至.渤海赤潮灾害监测与评估研究文集[M].北京:海洋出版社,2000:45-48.
[32] 国家海洋局.2009年中国海洋灾害公报[R].2009.
[33] 田家怡,闫永利,李建庆,等.山东海洋外来入侵生物与防控对策[J].海洋湖沼通报,2009(1):41-46.
[34] 邹景忠.海洋环境科学[M].济南:山东教育出版社,2004:284-300.
[35] 梁玉波,王斌.中国外来海洋生物及其影响[J].生物多样性,2001,9(4):458-465.
[36] 武正军,王彦平,李义明.浙江东部牛蛙的自然种群及潜在危害[J].生物多样性,2004,12(4):441-446.
[37] 赵建东.防止外来海洋物种入侵应多管齐下[N].中国海洋报,2009-05-26(A1).
[38] LIAO C Z,LUO Y Q,JIANG L F,et al.Invasion of Spartina alterniflora enhanced ecosystem carbon and nitrogen stocks in the Yangtze Estuary,China[J].Ecosystems,2007,10(8):1351-1361.
[39] 钦佩,谢民,陈素玲,等.苏北滨海废黄河口互花米草人工植被贮能动态[J].南京大学学报,1994,30(3):488-493.
[40] 朱洪光,陈小华,唐集兴,等.以互花米草为原料生产沼气的初步研究[J].中国沼气,2007,25(3):19-22.
[41] 清华,姚懿函,李红丽,等.互花米草生物质能利用潜力[J].生态学杂志,2008,27(7):1216-1220.
[42] 黄锡生.论我国防治外来物种入侵的法律对策[J].兰州大学学报,2005,33(1):109-114.
[43] 万方浩,郭建英,张峰,等.中国生物入侵研究[M].北京:科学出版社,2009:280-289.
我国近海海洋综合调查与评价专项(ZJ908-02-03-5、ZJ908-02-02、908-02-04-02);国家海洋局第二海洋研究所基本科研业务费专项(JT0806).
猜你喜欢
海洋通报(2022年5期)2022-11-30
中外文摘(2022年15期)2022-11-15
小哥白尼(趣味科学)(2022年7期)2022-09-20
农业灾害研究(2022年7期)2022-09-08
安徽农业科学(2022年14期)2022-08-04
海洋通报(2021年5期)2021-12-21
小天使·一年级语数英综合(2020年6期)2020-12-16
幽默大师(2019年11期)2019-11-23
中学生物学(2019年5期)2019-10-20
都市家教·下半月(2016年11期)2016-12-29
