
中等强度有氧运动大鼠环境应激的保护效应及其机制探讨
2012-10-13 05:02:58苏铁柱
长治学院学报 2012年5期
赵 振,苏铁柱
(长治学院 体育系,山西 长治 046011)
1 研究目的
当压力增大时,人们对突发事件的处理能力降低、对环境的适应能力变差,这种应激是目前多种疾病的病因和诱因[1]。研究发现,应激可引起一系列的脑功能障碍,如出现认知功能低下、情绪和行为异常等。此外,应激刺激还会诱发多种行为和状态的改变,如出现焦虑和抑郁状态、探究和攻击行为的改变、睡眠生物节律及活动量也有所影响等[2]。机体可通过激活神经内分泌系统的下丘脑-垂体-肾上腺皮质轴(hypothalamic-pituitary adrenalaxis,HPA)和交感神经系统(sympathetic nervous system,SNS)两条信号通路对应激反应产生调控。本实验通过观察大鼠运动训练后对外界损害性应激的各种行为学变化,并测定相关神经内分泌指标的改变,探讨中等强度有氧运动对大鼠应对外界损害性应激能力的影响及其机制,为人们通过运动锻炼增强应对突发事件及各种环境应激适应能力提供理论依据。
2 材料与方法
2.1 实验动物分组
三月龄健康雄性SD大鼠32只,体重180~220 g(购于西安交通大学医学院实验动物中心),大鼠适应性喂养1 wk,随机分为安静对照组(A组)、运动训练组(B组)、应激组(C组)、运动训练应激组(D组),每组8只。分笼饲养,每笼5只。均以国家啮齿类标准动物饲料喂养,自由饮水,饲养环境28℃±1℃,相对湿度为50-65%。
2.2 运动训练方案
B和D组大鼠运动训练采用递增强度的方式进行跑台运动,运动负荷参照Bedford等人的研究进行[3]。起始速度为11 m/min,时间为20 min,每周训练5天,递增速度3 m/min、时间5 min,运动至速度为20 m/min,维持此运动速度运动60 min,跑台坡度为5度,训练周期为8周。
2.3 应激实验方案
D组运动训练8 w后与C组大鼠一同接受电击、强声、强光多种损害性应激1 w[4]。将大鼠分别置于封闭的笼中,给予足底电击(电流强度1 mA、电压 30 V,每隔1 min刺激1次,每次持续10 s,单位电击周期为5 min);强噪声刺激(200 HZ强声刺激,每隔30 min刺激1次);强光刺激(500 w普通灯泡夜间照射8 h)。实验中应掌握同种刺激不要连续出现,使大鼠不能预料或习服刺激的发生。
2.4 旷场行为学观察
C、D组末应激后24 h,各组大鼠依次进行旷场实验,室内保持安静,光照度为60 Lux,噪音控制在65 dB以下。实验装置由旷场反应箱和数据自动采集处理系统两部分组成。旷场反应箱高30~40 cm,底边长50 cm,底部涂黑并将其平均分为25个4 cm*4 cm的小方格,正上方1 m处架一数码摄像头,其视野可覆盖整个矿场内部。实验时将实验动物置入旷场中央格内,通过行为学摄像系统记录动物3 min的行为,观察大鼠在中央格停留时间、穿越格数(四爪均进入的方格方可计数,为水平运动得分)、后肢站立次数(两前爪腾空或攀附墙壁,为垂直运动得分)、修饰次数和粪便粒数。每只动物仅做1次行为测定,用75%酒精彻底清洁敞箱后再进行下一只动物的观察[5]。实验期间密切观察各组大鼠一般行为学变化。
2.5 生化指标与测试方法
血浆皮质醇、去甲肾上腺素和多巴胺水平以及血浆NO和NOS含量的测试方法(采用南京建成生物工程研究所的试剂盒),严格按照试剂盒操作说明进行。
2.6 数据处理
实验数据均采用SPSS17.0统计软件进行处理,统计学方法采用One-Way ANOVA方差分析,实验结果均以平均数±标准差(±S)表示,显著性差异选择P<0.05和P<0.01水平。
3 实验结果
3.1 各组大鼠一般行为学变化
正常对照组大鼠生活状态正常,皮毛整洁光滑,对新异刺激及周围环境的变化反应灵敏,在笼内很活跃,进食较多,运动能力较强,探究性行为频繁。运动训练组大鼠较正常对照组显得更为活跃,食量增加,捕捉时逃避行为明显,一般状况正常,随着时间的推移,大鼠对外界刺激反应迟钝,探究性行为减少。应激组大鼠在应激后,躁动不安、高度警惕、活动能力显著下降,体重减轻,皮毛散乱、直立、无光泽,饮水、进食有所减少,多数静卧少动。而运动训练应激组大鼠较应激组相比,上述惊恐表现明显降低,能很好的应对损害性应激。
3.2 各组大鼠旷场行为学变化
与安静对照组相比,运动训练组大鼠穿越格数和站立次数均显著增加(P<0.01)。而应激组大鼠场中的潜伏期显著延长(P<0.01),其穿越格数、站立次数和修饰次数均显著减少(P<0.01),粪便粒数显著增多(P<0.05)。与应激组相比,运动训练应激组大鼠场中潜伏期显著缩短(P<0.01),穿越格数和修饰次数均显著增加(P<0.01),站立次数和粪便颗粒显著增加(P<0.05)。

表1:各组大鼠旷场行为相关指标数据统计
3.2 血浆皮质醇、去甲肾上腺素和多巴胺水平的变化
与安静对照组相比,运动训练组大鼠血浆皮质醇、去甲肾上腺素和多巴胺水平均显著增加(P<0.05)。应激组大鼠血浆皮质醇水平显著增加(P<0.01),去甲肾上腺素水平显著降低(P<0.05),多巴胺水平则极显著降低(P<0.01)。与应激组相比,运动训练应激组大鼠多巴胺水平显著增加(P<0.01),去甲肾上腺素也显著增加(P<0.05)。

表2:各组大鼠血浆皮质醇、去甲肾上腺素及多巴胺水平的变化
3.3 各组大鼠血浆NOS-NO含量变化
与安静对照组相比,运动训练组大鼠血浆NO和肝脏NOS含量均显著增加(P<0.01);应激组大鼠血浆NO含量显著增加(P<0.01)。与应激组相比,运动训练应激组大鼠肝脏NOS含量显著增加(P<0.01),NO含量也显著增加(P<0.05)。
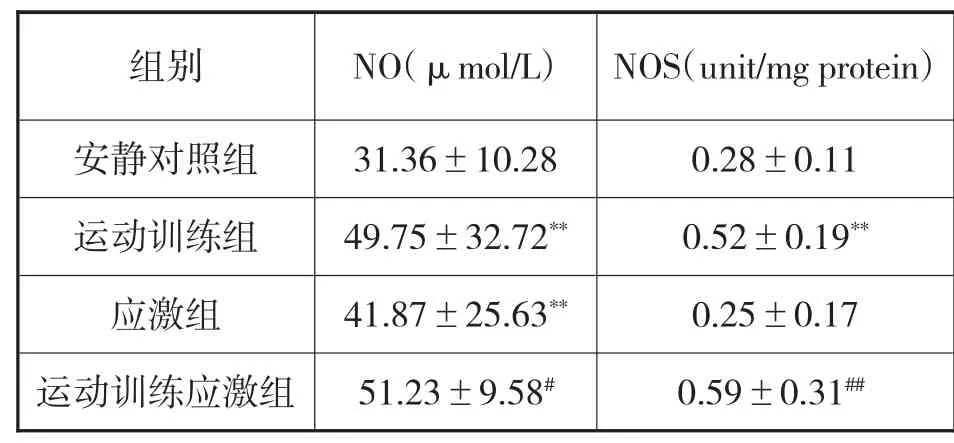
表3:各组大鼠血浆NOS-NO含量变化
4 分析讨论
近年来,有关应激对机体损伤作用的研究越来越多,人们对该问题的重视程度越来越高。应激是机体对各种应激源的非特异性适应反应,各种引发应激反应的刺激反应又可以分为躯体性应激(如热刺激、冷刺激和疼痛刺激等)和心理性应激(如考试),一般情况下两者同时发生,相互混杂[6]。心理性应激涉及到生理和行为方面的变化,互相之间具有密切的联系。行为方面主要表现为攻击、退缩等。在生理方面,往往引起较大的认知或生理上的反应,使交感神经活动增强、副交感神经活动减弱,并使机体免疫功能受到抑制,这就要求机体作出适应性的反应来保持其内部的稳定性[7]。大量研究表明,当一些心理应激情况出现时,其神经内分泌免疫调节网络、行为能力和学习记忆能力都会受到明显影响[8]。此外,许多研究也显示:适量的运动可以使糖皮质激素的分泌减少,对皮质醇的释放有良好的调节作用,能抗糖皮质类激素的过量分泌[9]。
4.1 中等强度运动训练后大鼠对应激刺激的一般行为的影响
应激对大鼠的食量、活动能力、反应力等一般状况等都会产生一定的影响。本实验研究结果显示:与正常对照组相比,给予大鼠强损害性应激后,应激组大鼠极度恐慌,发生紊乱、慌乱的逃避行为,或蜷缩不动的僵持状态,有的甚至发生自残、相互残杀等现象;而与运动训练组相比,运动训练应激组大鼠的恐慌表现显著降低,表明强烈的损害性应激使得机体正常免疫监控失调,造成行为和认知的改变。通过早期进行运动训练干预,不仅能够有效降低应激损害,增强机体对抗应激损害的能力及环境适应能力,同时还可有效维持机体的正常功能和保持其内部的稳定性。
4.2 中等强度运动训练后大鼠对应激刺激的旷场行为的影响
旷场实验(Open-field test)又称敞箱实验,是以实验动物在新奇环境中某些行为的发生频率和持续时间为标准,对动物的自主行为和探究行为进行评价,以尿便次数来判断其紧张度的一种应激行为学研究方法[10]。其中穿越格数及站立次数是反应动物探索行为及兴奋性的指标;中央格时间反映动物对环境的认知能力。正常动物会避开空旷环境,迅速离开中央格,沿周边活动。如果对新环境的认知能力差,则停留在中央格的时间就会延长;修饰是动物满足和自我关注的表现;排便多反映了动物的紧张强度高。
本实验研究结果显示:与安静对照组相比,运动训练组大鼠穿越格数和站立次数均显著增加(P<0.01),表明运动训练能够使得大鼠探索欲增加,兴奋性提高,能够很好的适应新环境;而应激组大鼠在场中的潜伏期显著延长(P<0.01),其穿越格数、站立次数和修饰次数均显著减少(P<0.01),粪便粒数显著增多(P<0.05)。表明应激使大鼠的探索行为下降,兴奋性显著降低,对新环境的适应性和对空间的认知能力均比正常动物差,且表现出较高的紧张度。与应激组相比,运动训练应激组大鼠场中潜伏期显著缩短(P<0.01),穿越格数和修饰次数均显著增加(P<0.01),站立次数和粪便颗粒显著增加(P<0.05),运动训练应激组大鼠在探索行为及兴奋性,对新环境的适应性比单纯应激组要强很多,而在紧张度上明显比单纯应激组低,表明运动训练干预大鼠后对其应对外界应激刺激具有显著缓解与适应作用。
4.3 中等强度运动训练后大鼠对应激刺激行为学变化的可能机制分析
应激反应的主要特征是神经内分泌反应增强,其中以HPA轴终端肾上腺皮质分泌的糖皮质激素(主要是皮质醇)增高为显著特征。此外,SNS激活后可从交感神经末梢和肾上腺髓质释放儿茶酚胺,随着应激强度的逐渐增加,糖皮质激素和儿茶酚胺的分泌呈增加趋势[11]。
皮质醇是人体内主要的糖皮质激素,由肾上腺皮质束状带分泌,其分泌与合成受下丘脑-垂体-肾上腺皮质轴调节,血浆皮质醇水平具有明显昼夜节律。研究发现,应激刺激时,下丘脑分泌的促肾上腺皮质激素释放因子,可刺激促肾上腺皮激素释放,后者作用于肾上腺皮质能够促进皮质醇分泌,皮质醇分泌反过来又可抑制促肾上腺皮激素的释放。损害性应激可明显引发大鼠的不良情绪反应,增强肾上腺皮质激素的分泌,导致皮质醇含量升高。有研究表明,皮质醇变化与运动强度和运动量之间有一定的量变关系,适度的身体锻炼对皮质醇的释放有良性的调节作用[12]。本研究发现,与安静对照组相比,应激组大鼠血浆皮质醇水平显著增加(P<0.01),而运动训练组大鼠血浆皮质醇水平均显著增加(P<0.05);与应激组相比,运动训练应激组大鼠血浆皮质醇水平有所降低,但无显著差异,说明血浆皮质醇水平改变与行为学变化具有相关性。本研究结果表明,适度的运动训练可通过降低应激引起的皮质醇水平进而缓解大鼠行为功能减弱状况。
去甲肾上腺素和多巴胺是SNS激活后从交感神经末梢和肾上腺髓质释放儿茶酚胺类递质,主要参与调节机体的觉醒、注意力和精神情绪状态。应激水平与血浆去甲肾上腺素浓度增加相关[13];多巴胺在黑质纹状体中含量丰富,参与机体认知过程,并与精神活动密切相关。其精神情绪状态则主要表现在行为学自主活动及对外界环境应激的适应性。应激诱导的行为异常与去甲肾上腺素和多巴胺水平耗竭有关。在应激情况下,多巴胺神经元生理活动加强,当应激的强度或持续时间超过机体耐受能力时,将导致认知功能及行为能力的下降。此时,酪氨酸的充足供应能增强多巴胺的合成和释放,能在一定程度上拮抗应激对中枢多巴胺神经元的损害[14]。酪氨酸属于中性芳香族氨基酸,是合成儿茶酚胺类神经递质的前体物。运动训练可能促进酪氨酸生成以适应应激刺激及外界环境。本研究结果显示,与安静对照组相比,运动训练组大鼠去甲肾上腺素和多巴胺水平均显著增加(P<0.05),应激组大鼠去甲肾上腺素水平显著降低(P<0.05),多巴胺水平则极显著降低(P<0.01)。与应激组相比,运动训练应激组大鼠多巴胺水平显著增加(P<0.01),去甲肾上腺素也显著增加(P<0.05)。表明中等强度有氧运动可通过增加儿茶酚胺类递质的释放应对应激反应,产生保护效应。
NO在神经系统内作为神经递质不仅具有信息传递功能,而且具有拮抗谷氨酸介导的神经毒性作用,该作用可能是通过阻断NMDA受体与谷氨酸结合实现的[15]。实验显示,运动训练应激组NOS及NO含量升高,推测运动训练可能具有增强机体NOS-NO水平作用,从而提高大鼠的应激适应性,其可能机制还需更深入的研究。
综上,本研究发现应激刺激引起大鼠旷场行为改变,表现为中央格停留时间延长,穿越格数、站立次数和修饰次数均显著较少,粪便颗粒增多,应激激素分泌增加,而预先经运动训练大鼠这种由应激造成的不良反应明显减轻,其机制可能是通过改变神经递质去甲肾上腺素和多巴胺水平的变化及机体NOS-NO水平实现。
[1]WR Lovallo.Stress and health:Biological and psychological interactions[M].London:Sage Publications,2005.
[2]王玮文,邵枫,林文娟.不同时程应激对大鼠行为、免疫和交感神经系统反应的影响[J].心理学报,2007,39(2):292-298.
[3]Bedford TG,Tipton CM,Wilson NC,et al.Maximum oxygen consumption of rats and its changes with various experimental procedures[J].J Appl Physiol,1979,47(6):1278-1283.
[4]张莉,崔伟,黄昌林.模拟心理训练对大鼠机体应激能力的影响[J].解放军医学杂志,2007,32(12):1240-1242.
[5]魏宏文,矫玮,张有志等.运动对慢性应激抑郁模型大鼠行为学及体重的影响[J].体育学刊,2010,17(11):100-105.
[6]宋淑玲,王君明.MSG对大鼠学习记忆能力及旷场行为影响的实验研究[J].广州医药,2008,39(1):1-4.
[7]张莉,崔伟,黄昌林.心理训练对应激后大鼠旷场行为的影响[J].实用医药杂,2008,25(8):989-990.
[8]de Quervain DJ,Roozendaal B,McGaugh JL.Stress and glucocorticiods impair retrieval of long-term spatial memory[J].Nature,1998,394(6695):787-790.
[9]路翠艳,潘芳.应激反应中HPA轴的中枢调控和免疫调节[J].中国行为医学科学,2003,(12):353-355.
[10]王雪琦,孙学军等.睫状神经营养因子对皮质酮致海马神经元损伤的保护作用[J].第二军医大学学报,2000,21(5):406-408.
[11]Lawrence DA,Kim D.Central/peripheral nervous system and immune response[J].Toxicology,2000,142(3):189.
[12]潘纪戌,吴国庚,陈起航等.肺气肿的诊断:CT和肺功能检查的比较[J].中华放射学杂志,1995,(10):680-683.
[13]Fujiwara K,Tsukishima E,Kasai S,et al.Urinary catecholamines and salivary cortisol on workdays and days off in relation to job strain among:female health care providers[J].Scand J Work Environ Health,2004,30(2):129-138.
[14]陈伟强,程义勇,李树田等.心理应激对大鼠旷场行为的影响及酪氮酸干预作用研究[J].中国应用生理学杂志,2009,24(1):125-127.
[15]李凌江,李则宣.精神应激的生物学致病机制(Ⅱ):神经内分泌机制[J].中国行为医学科学,2005,14(2):156-158.
猜你喜欢
中国急救医学(2022年2期)2022-11-15 09:04:10
中国典型病例大全(2022年12期)2022-05-13 14:57:02
医药导报(2021年10期)2021-10-09 04:21:34
疯狂英语·新读写(2020年3期)2020-06-06 09:05:54
中国比较医学杂志(2018年9期)2018-01-24 09:05:24
现代养生·下半月(2017年7期)2017-12-28 13:09:52
中国体育教练员(2017年2期)2017-07-31 19:09:34
合成化学(2015年10期)2016-01-17 08:56:07
中国民族民间医药·下半月(2014年2期)2014-09-26 05:21:22
安徽医药(2014年5期)2014-07-07 15:29:39
