
夏季高温下猕猴桃不同节位叶片光合作用研究
2012-10-10 06:48何科佳李健权卜范文王中炎
湖南农业科学 2012年21期
何科佳,李健权,杨 玉,卜范文,曾 斌,王中炎
(1.湖南省园艺研究所,湖南 长沙 410125;2.国家农业部华中地区果树科学观测站,湖南 长沙 410125;3.长沙时鲜水果工程技术研究中心,湖南 长沙 410125)
猕猴桃为落叶藤本果树,属于猕猴桃科(Actini diaceae)猕猴桃属(Actinidia Lindl.)植物,是一种新兴的营养保健水果。从上世纪80年代开始,猕猴桃在全球范围内进入飞速发展时期。随着猕猴桃产业的飞速发展,猕猴桃科研工作也全面开展。猕猴桃光合作用的研究也取得了不少成绩。猕猴桃不同品种、不同气候条件、不同环境因子对猕猴桃光合作用的影响及其生理、分子基础都做了一定研究[1-3]。同一植株或同一枝条上不同节位叶片光合特性的差异,可能在很大程度上反映了叶片生长发育时期或叶龄的差异,同时也可能在一定程度上受到生长期间微环境条件差异的影响[4-5]。因此,在测定活体条件下,猕猴桃不同节位叶片的光合特性可以更真实地了解自然条件下不同发育时期叶片的光合特性。该研究主要探讨在低海拔地区,夏季高温条件下,不同节位猕猴桃叶片的光合作用特点,为猕猴桃向低海拔、夏季高温地区发展提供基础数据参考。
1 材料与方法
1.1 试验地点
试验地点在湖南省园艺研究所猕猴桃试验示范基地。该基地位于长沙市东郊(海拔50 m),夏季最高气温一般在40℃左右,且常伴随严重干旱,极具代表性,是理想的试验场所。
1.2 试验材料
试验材料为8年生中华猕猴桃中熟品种翠玉,10月上旬成熟,采用大棚架栽培,生长势中等,夏季因高温常出现落叶落果现象。
1.3 试验方法
选晴朗无云的天气,于 8∶00、10∶00、12∶00、14∶00、16∶00、18∶00 用 CI-310 型便携式光合测定系统测定,每个时间点测6个数据求平均值。
供试叶片选择:从供试植株上部选择不同方位且光照条件良好的3个结果母枝作为供试枝条,从供试枝条上的基部(1~3节),中部(6~10节),梢部(倒数1~3节)选择一片完整,健康能满足叶室要求的叶片进行测定。
1.4 数据分析
所有数据均采用Excel2003和SPSS11.5数据分析软件进行处理和分析。
2 结果与分析
2.1 猕猴桃不同节位连体叶片叶幕微环境分析
叶片有效光合辐射(PAR)如图1所示,不同节位叶片的PAR日变化趋势基本一致,呈单峰曲线型,峰值均出现在午间13∶00左右,早晚较低,变化幅度在检测时间内为200~1 900 mol/(m2·s)。但不同节位叶片由于受到周围叶片的影响,全天的光照条件有一定的差异,基部叶片<中部叶片<梢部叶片。基部叶片由于处于植株中央位置,最为郁闭,而梢部叶片一般分布于植株外围,光照最强,特别是在午间强光时段,差异最为明显。但供试叶片均处于夜幕上部,光照条件较好,因此PAR全天都无显著性差异。

叶幕温度(Tair):不同节位叶片Tair日变化趋势基本一致,呈单峰曲线型,早晨最低,之后几乎直线攀升,13∶00左右达到最高值,超过38℃,瞬时极端最高气温超过40℃。在13∶00~15∶00气温相对稳定,变幅不大,均保持在37℃以上。15∶00之后开始明显下降(图2),下午17:00左右降到约34℃。Tair与PAR的变化趋势基本一致。不同节位叶片Tair存在一定差异,但不显著。

叶幕空气相对湿度(RH):不同节位叶片RH日变化趋势也基本一致,呈“V”型。早晨最高,超过80%;然后逐渐下降,15∶00左右出现最低值(50%以下)后又稳步回升(图3),17∶00接近60%。就全天的RH而言,梢部叶片的相对湿度略低于中部、基部叶片,但均无显著性差异。

叶幕CO2浓度(Ca):基部与中部叶片Ca日变化趋势基本一致,呈“V”型变化,早晚较高,午间较低(图4)。但基部、中部叶片的最低值出现在11∶00左右,13∶00后迅速回升;而梢部叶片的最低值出现时间较基部、中部的有所延后,出现在13∶00左右,之后迅速回升。梢部叶片Ca也略高于基部和中部,这可能是由于梢部处于外围、比较通风的缘故,但3者间也均无显著性差异。综合以上分析,猕猴桃不同节位叶片叶幕微气候存在一定差异,但在PAR、RH、Tair和Ca上均无显著性差异。

2.2 猕猴桃不同节位叶片净光合速率分析
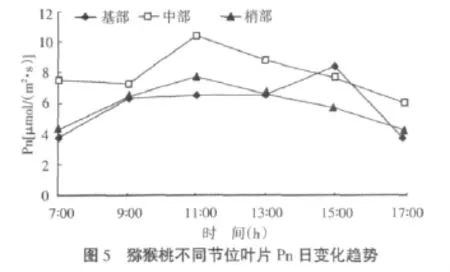
如图5所示,中部与梢部叶片的净光合速率(Pn)日变化趋势均为单峰曲线型,峰值出现在11∶00左右,之后随着环境条件的变化(高温、强光),Pn迅速下降,15∶00以后在高温、强光胁迫有所缓解的情况下也未出现Pn的回升。梢部叶片也呈单峰曲线型,但峰值出现在15∶00左右,之后迅速下降。另有数据显示:基部(5.95)和梢部(5.91)叶片Pn的全天平均值无显著性差异,但大大低于中部叶片(8.01),存在显著性差异。
从图6中可以看出,不同节位叶片气孔导度(Gs)的变化规律基本一致,13∶00之前均处于低位,在10 mmol/(m2·s)以下;13∶00之后迅速上升,17∶00到达最大值。15∶00之前,不同节位叶片的Gs很接近,15∶00之后差异拉大,梢部叶片>中部叶片>基部叶片。从图7中可以看出,不同节位叶片Ci日变化趋势极为相似,均呈“U”型曲线变化,最低值出现在11∶00 左右。

3 小结与讨论
高温对猕猴桃叶片的光合作用有很大的影响[6-8]。在高温强光胁迫下,基部、中部、梢部叶片的Pn日变化趋势都呈单峰曲线型,中部与梢部叶片的Pnmax均出现在上午11∶00左右,而基部叶片的峰值则出现在15∶00左右,叶肉细胞光合活性的降低很可能是高温强光胁迫下Pn下降的主要原因。猕猴桃不同节位叶片的Pn存在显著性差异:梢部叶片由于受到发育状况的影响,自身的光合能力较弱,加之处于环境胁迫最严峻的树冠外围,其Pn显著低于中部叶片;而因为老化和处在较为郁闭的冠幕微环境下,基部叶片Pn与中部叶片也存在显著性差异。综合分析发现,中部叶片11∶00之后Pn的下降与Ca(图4)、Gs(图6)和Ci(图7)的上升几乎同步发生,由此可见在高温强光胁迫下,叶肉细胞光合活性降低很可能是Pn下降的主要原因。但是气孔导度(Gs)的上升并不意味着可以排除气孔限制的因素,因为它也可能是气孔部分关闭造成的假相。基部连体叶片的Pn日变化也基本上呈单峰曲线型,但其最高值出现在15∶00左右,这可能是因为基部叶片处在光照相对低一些的树冠内部,所受到的胁迫较小,且下午气孔导度有所增加。
研究发现,长沙作为低海拔、高温地区,盛夏高温对叶片全天的净光合速率有一定抑制作用。类似地区要发展优质猕猴桃需采用一定的栽培措施,如适度遮荫、加强水肥管理增强树势等。
[1]何科佳,王中炎,王仁才.夏季遮荫对猕猴桃园生态因子和光合作用的影响[J].果树学报,2007,(5):50-53.
[2]何科佳,王中炎,王仁才.夏季遮荫对猕猴桃生长发育的影响[J].湖南农业科学,2007,(1):41~43.
[3]何科佳,王中炎,王仁才.高温干旱强光对猕猴桃生长发育的影响及其生理基础[J].湖南农业科学,2005,(3):42-44.
[4]袁飞荣,王中炎,卜范文,等.夏季遮荫调控高温强光对猕猴桃生长与结果的影响[J].中国南方果树,2005,34(6):54-56.
[5]耶兴元,马锋旺,王顺才,等.高温胁迫对猕猴桃幼苗叶片某些生理效应的影响[J].西北农林科技大学学报(自然科学版),2004,32(12):33-37.
[6]沈维良,柳建国.作物耐旱性与叶片水势简报[J].中国生态农业学报,2001,9(4):100-101.
[7]谢志兵,杨清平,黄云霞,等.猕猴桃叶片光合特性初探[J].林业科学,2003,28(3):49-50.
[8]姜卫兵,高光林,俞开锦,等.水分胁迫对果树光合作用及同化代谢的影响研究进展[J].果树学报,2002,19(6):416-420.
猜你喜欢
塔里木大学学报(2021年1期)2021-03-31
长江丛刊(2020年13期)2020-11-19
数学物理学报(2020年2期)2020-06-02
蔬菜(2020年1期)2020-01-17
婚姻与家庭·婚姻情感版(2019年6期)2019-06-03
青少年科技博览(中学版)(2018年6期)2018-08-16
中外葡萄与葡萄酒(2017年3期)2017-06-09
合作经济与科技(2015年22期)2015-08-09
小小说月刊·下半月(2012年12期)2012-05-14
长江蔬菜(2011年2期)2011-03-22
