
翠柏木材解剖性质和结晶度的径向变异及化学性质1)
2012-09-18 01:27崔凯
东北林业大学学报 2012年4期
崔 凯
(中国林业科学研究院资源昆虫研究所,昆明,650224)
孙庆丰
(东北林业大学)
廖声熙
(中国林业科学研究院资源昆虫研究所)
石江涛
(东北林业大学)
王小庆 李 昆
(中国林业科学研究院资源昆虫研究所)
目前全球用材资源逐渐从天然林转向人工林,大力发展人工林已是世界各国面对天然林和天然次生林日益减少所采取的共同战略。许多工业化国家和发展中国家都把大力发展人工林作为解决二十一世纪木材需求的根本措施,并制定了长期的人工林发展规划,以此来解决生态环境和木材供需之间的矛盾。人工林已经成为世界木材供应和木材加工利用原料的主要来源,因此,人工林已成为当代世界木材科学的主要研究对象[1]。近年来,人们已经认识到“林木—木材材性—加工利用”是一个有机的整体,木材材性既是人们对林木实行定向培育的技术依据,也是评价人工用材林营造成功与否的关键[2]。
翠柏(Calocedrus macrolepis Kurz)系柏科翠柏属常绿乔木,国家Ⅱ级重点保护野生植物,全球仅有一种,属于古老残遗物种[3]。陈文红[4]、宁世江[5]等对翠柏的生物群落特征研究表明翠柏群落是相对稳定性较大的群落,对其研究可为开发利用该木材及对气候生态的演变提供重要的信息。陈子牛[6]、谭开湛[7]、张茂钦[8]等从生态、濒危珍稀林木资源保护和开发方面对翠柏做了调查研究。吕玉华[9]利用野生翠柏苗进行人工培育翠柏小苗以期进行大面积培育该树种。张学星[10-11]等对翠柏的抗污染气体和净化空气的性能做过研究,认为翠柏是一种可有效净化空气的优良树种。其它虽仍有一些有关翠柏木材的相关研究[12-15],但由于其分布的特异性,国外对其材性方面的研究比较少,而对翠柏木材材性系统分析的文献资料更为少见。本研究对云南产的翠柏木材解剖性质、化学性质和结晶度的径向变异做出分析,对解剖各项参数的特征和结晶度间关系建立预测模型,为了解翠柏木材材性的变异和加工利用提供基础依据。
1 材料与方法
试材采自云南省昌宁县漭水镇,选取翠柏分布较为集中的区域,从中选取树干通直,生长正常,态势均一的样木5株,树龄52年生。采集时间2009年7月,采集方法依据中华人民共和国国家标准木材物理力学性质试验方法(GB1927—91)[16]中所述方法进行。样木选定后,标出南北向后进行砍伐,伐倒后即在每株的胸径处截取1个30 mm厚圆盘运回实验室放置风干后待用,每个圆盘沿东西方向截取宽4 cm的中心试条,沿着径向按每5生长轮(共取测样点10个)由髓心向外取样,取出的样品进行解剖性质、化学性质和结晶度测定。
解剖性质试验:在伐倒后的木材树干1.3 m处向上连续截取两个厚约2 cm的圆盘,其中一个圆盘按南北方向过髓心取1.5 cm宽的中心木条,用于测定解剖参数,利用常规方法切片、离析与测定[17],微纤丝角测量采用碘结晶法。
结晶度试验:另外一个圆盘按上述方法取下木条后,按生长年轮从髓心到树皮方向分别取下各个生长年轮木材,每株树中每生长轮中取4个试样测定木材结晶度。测定前将试样磨成木粉,取80~100目颗粒的木粉。采用D/MAX2200VPC型X射线衍射仪,日本理学制造,X光管为铜靶,管电压为40 kV,管电流为35 mA,扫描步距为 0.02°,扫描范围为10°~40°,扫描速度为 4°/min,木粉压片后进行扫描,采用Segal法计算结晶度[18]。
化学性质试验:在每根原木梢部、中部和基部分别截取3个10 cm厚的圆盘,剥皮并去除有缺陷部分,切成小薄片,采用四分法充分混合均匀后磨成木粉,过筛,取通过40目而留于60目的部分作为化学成分含量分析试样。化学成分的测定按照GB/T2677.2-5—93 及 GB/T2677.6-9—94[19]所规定的方法进行,其中纤维素质量分数的测定采用硝酸―乙醇法[20],木材 pH 值的测试方法按照 GB6043-85[21]中所规定的方法。
使用Excel2003、SPSS16.0对所得数据进行处理、分析和模型建立。
2 结果与分析
2.1 解剖性质和结晶度的径向变异模式和变异
2.1.1 管胞形态的径向变异模式
翠柏木材每5生长轮的管胞形态特征值的径向变异曲线如图1和图2所示。管胞长度、宽度和壁厚的径向变化趋势基本一致,从髓心向外增加,木材生长20 a以前增加趋势十分明显,20 a之后增加趋势减缓,说明20~25 a翠柏木材尚处于树木生长的旺盛期,形成层细胞分裂活动能力较强,反映到管胞形态上就是各形态特征值迅速增大。当翠柏木材进入成熟期后,形成层细胞分生能力趋于稳定,反映在管胞形态的变异曲线上就是各管胞形态特征值的变异趋势也趋于稳定。翠柏木材的管胞长度从1886 μm 增加到 3370 μm,与 Panshin[22]等关于针叶材管胞长度的径向变化曲线的论述一致,为PanshinⅡ型曲线。纤维宽度从11 μm增到20 μm,管胞壁厚的变化范围是3.1~9.3 μm。其中,管胞长度的变异曲线波动较大,宽度和壁厚的变异曲线略平缓,各生长轮间管胞形态差异非常明显。采用对数模型建立管胞长度、宽度及壁厚径向变异回归模型。其中,管胞长度的回归模型是y=679.9lnx+1961,管胞宽度的回归模型是y=4.484lnx+9.77,管胞壁厚的回归模型是 y=2.364lnx+4.124,R2分别是 0.8855、0.9733和0.8657,相关性非常显著。
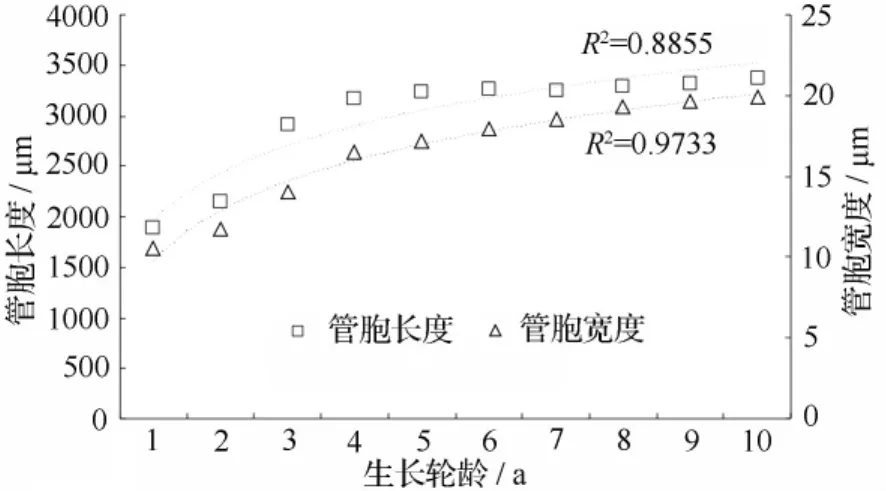
图1 管胞形态径向变异曲线
2.1.2 组织比量的径向变异模式
木材组织比量是组成木材不同细胞组织所占木材的比例,主要表征木材显微组织构造的体积量特征。木材组织比量是决定木材物理力学性质的因素之一,与木材性质、木材的机械和化学加工性能以及利用密切相关,同时也为定向培育和材质预测提供重要的依据。翠柏木材各组织比量的径向变异曲线见图2和图3,可见,管胞比量的变化范围为89.23% ~90.57%,变幅为1.34%;射线比量的变化范围是8.92% ~10.51%,变幅是 1.59%;薄壁组织比量的变化范围是0.85% ~1.21%,变幅为0.36%,差异明显。各生长轮间组织比量的变异没有明显规律,首先采用对数模型建立生长轮龄和各组织比量间的回归模型,无法建立合适的变异趋势模型,然后采用三阶移动平均模型对各组织比量进行平滑处理得到的组织比量序列仍无法建立合适的变异趋势模型,说明木材组织比量径向变异生长趋势较小。
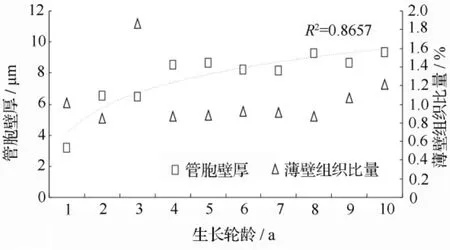
图2 管胞宽度和薄壁组织比量径向变异曲线
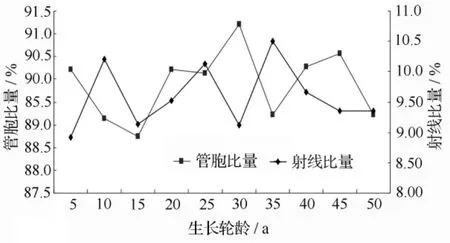
图3 管胞比量和射线比量径向变异曲线
2.1.3 微纤丝角和胞壁率的径向变异模式
胞壁率是指木材结构中除去细胞腔部分,组成木材实质部分所占的百分率,胞壁率对木材的力学性质有很大影响。翠柏木材各生长轮间胞壁率和微纤丝角的径向变异曲线如图4所示,胞壁率的变化范围是40.58% ~62.34%,变幅为 21.76%,变异极为显著;变异趋势和管胞形态的变异趋势基本相似,木材生长20 a之前变异较大,随后趋于稳定。翠柏木材微纤丝角的径向变化趋势是随着生长轮的增加而减小。大量研究已经表明,木材微纤丝角径向的变异一般模式为:在幼龄材区域由内向外降低,变异幅度一般在10°左右[23],本研究的结论和他们一致。微纤丝角的大小从根本上决定木材强度,微纤丝角越大,木材强度相对较低;反之,木材的强度就越高。该木材的微纤丝角平均值为14.21°,变化范围为18.35°~11.62°,变幅是 6.73°,差异非常显著。用对数模型分别对木材胞壁率、纤丝角和生长轮龄进行回归分析,胞壁率与生长轮龄间的回归模型是y=8.44ln(x)+44.58,纤丝角和生长轮龄间的回归模型是 y=-2.623ln(x)+18.17,R2值分别为 0.8742和0.928,相关性极其显著。

图4 微纤丝角和胞壁率径向变异曲线
2.1.4 结晶度的径向变异模式
木材结晶度是指木材中纤维素构成的结晶区占纤维素整体的百分数,与木材的生长特性、组织结构和化学性质有着密切关系,同时对木材尺寸稳定性、密度和硬度等具有重要的影响[24],一定程度上可以反映木材纤维的物理和化学性质。因此,研究木材的结晶度对于从组织构造和化学组成上分析木材材性具有指导意义。目前,常用来测定木材结晶度的方法就是X射线衍射法,简称为XRD。图5是翠柏木材的XRD图,可以看出,在2θ=22.8°附近的衍射峰最富于变化,这反映了该处衍射峰的大小与木材结晶度的大小变化具有一定的规律性,同时说明此晶格衍射峰与木材结晶度直接相关。
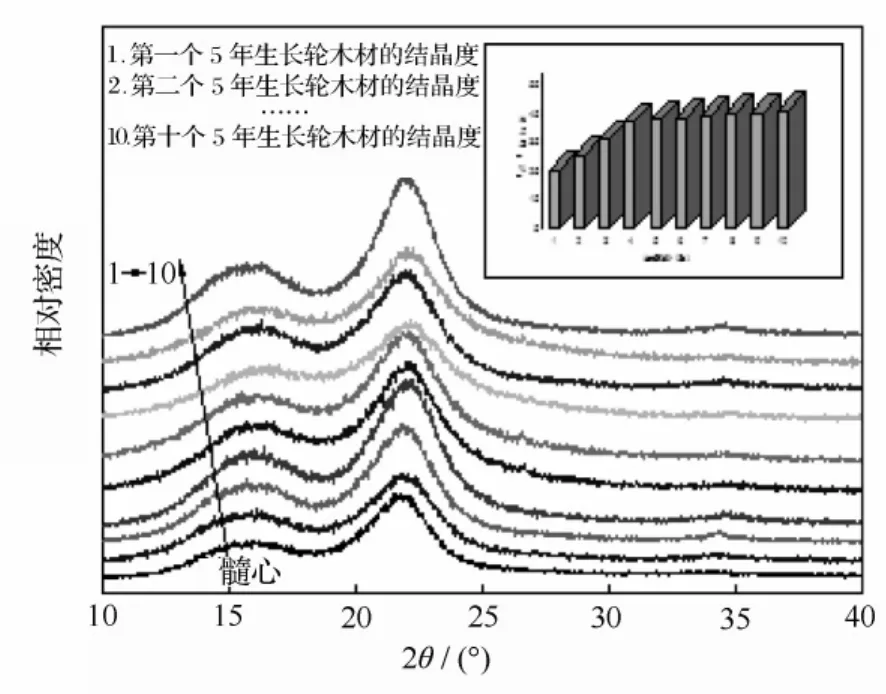
图5 木材各生长轮间的X射线衍射图
木材各生长轮间结晶度的径向变异趋势见图6。可以看出,各生长轮间木材结晶度的变化趋势和管胞形态及纤丝角和胞壁率的类似,木材生长20 a之前显著增加,之后变缓。各生长轮间木材的结晶度变化范围为25.3% ~45.7%,变幅为20.4%,差异非常显著。使用对数模型对结晶度与生长轮龄进行回归分析,回归模型是 y=9.381lnx+25.96,R2值为0.946,相关性非常显著。研究结果为利用木材结晶度随生长轮变化的趋势建立对木材结晶度预测模型提供了理论基础,这样不但可为及时调整林木培育和遗传改良措施提供参考,也可为人工林木材的高效利用提供科学依据。
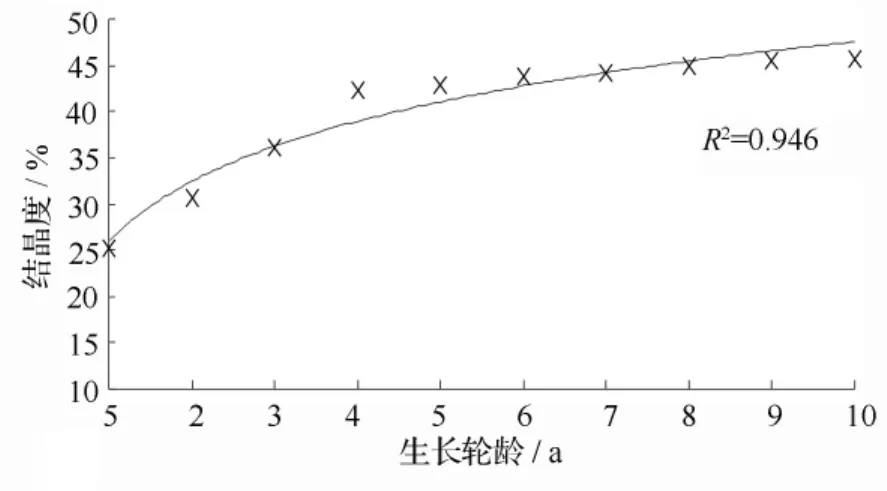
图6 结晶度径向变异曲线
2.2 各解剖参数和结晶度的相关性
2.2.1 结晶度与各解剖参数间相关性
木材各解剖参数和结晶度的相关分析结果见表1。木材结晶度与解剖参数间相关分析在石雷课题组[25]对印度黄檀(Dalbergia sissoo)的研究中有报道。笔者对翠柏木材结晶度与各解剖参数间的相关性进行探索性研究,以期为将来他们之间的相关性分析积累一些基础数据。对他们相关性的分析可为进一步了解木材性质和用少量的材料来研究木材性质打下一定的基础。从表1可以看出,木材的结晶度并不是与所有的解剖参数呈现相关性,说明并不是所有的解剖构造因子会对木材的结晶度产生影响,该研究结果和石雷课题组[25]相一致。结晶度与管胞长度、宽度、厚度和胞壁率均呈0.01水平显著正相关,与各组织比量无相关性,这也与上述对各组织比量的分析结果相一致,本研究中的翠柏木材的组织比量没有明显的变异规律。木材结晶度与纤丝角呈0.01水平显著负相关,说明木材的纤丝角随结晶度的增加而降低,与上述研究结果一致。
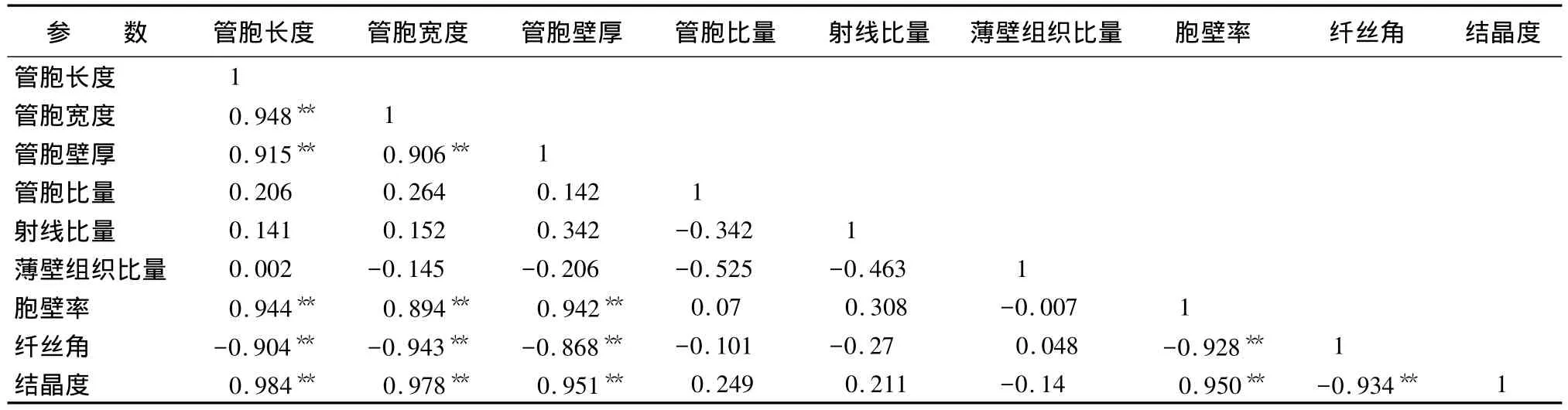
表1 各解剖参数和结晶度的相关分析
2.2.2 各解剖参数间的相关性
表1同时也对各解剖参数间的相关性进行了分析。可知,管胞各形态值间存在0.01水平上的显著正相关,胞壁率与管胞各形态值间呈0.01水平显著正相关,组织比量间及其与其它解剖参数间无任何相关性,纤丝角除与组织比量间无相关性外,与其它各解剖参数呈0.01水平上的显著负相关,说明大部分解剖参数都或多或少地对木材微纤丝角产生一定的影响;同时这些研究结果与上文中分析的木材各解剖参数的径向变异规律相符合。总之,通过对各解剖参数间相关性的分析可对各木材解剖性质及各解剖参数间复杂的关系有更深的了解,同时也为如何利用少量的解剖因子特征来分析其他解剖参数提供一定的基础数据。
2.3 结晶度与解剖参数(除组织比量外)变异模型的建立与预测
由于木材结晶度各解剖参数间存在着或正或负的显著相关,所以可对利用结晶度预测木材其它解剖参数进行探讨。模型的建立对木材材性的预测和减少试验样本使用量有着重要的意义。分析数据,使用Matlab语言编程,对产生的曲线进行拟合,最终建立木材解剖参数和结晶度间预测模型如表2所示。可以看出相关系数R2均在0.9以上,故此模型可用于本研究中的翠柏木材的解剖参数的评价、预测和验证,但如何建立更为广泛适用的预测模型仍需进行深入探讨。

表2 木材结晶度与各解剖参数的相关模型
2.4 化学性质
木材的主要化学成分有灰分、冷热水抽提物、1%NaOH抽提物、苯醇抽提物、半纤维素(阔叶材中以多戊聚糖为主)、木质素和纤维素等。纤维素是木材细胞壁的骨架物质,与木材性能、纸浆得率及纸浆质量关系密切。综纤维素是指除去半纤维素后剩余纤维素和木质素质量分数的总和,影响着木材的价值和使用范围。木质素除赋予木材强度外,还给予木材一定的尺寸稳定性。木材化学成分中抽出物质量分数影响着木材天然耐久性、材色和木材的加工利用。木材的pH值对制定木材应用范围和加工工艺具有重要意义,与木材胶合、木材变色、金属腐蚀和胶接性能等有密切关系。表3列出了翠柏木材各化学 组分,可以为翠柏木材最佳利用提供基础数据。
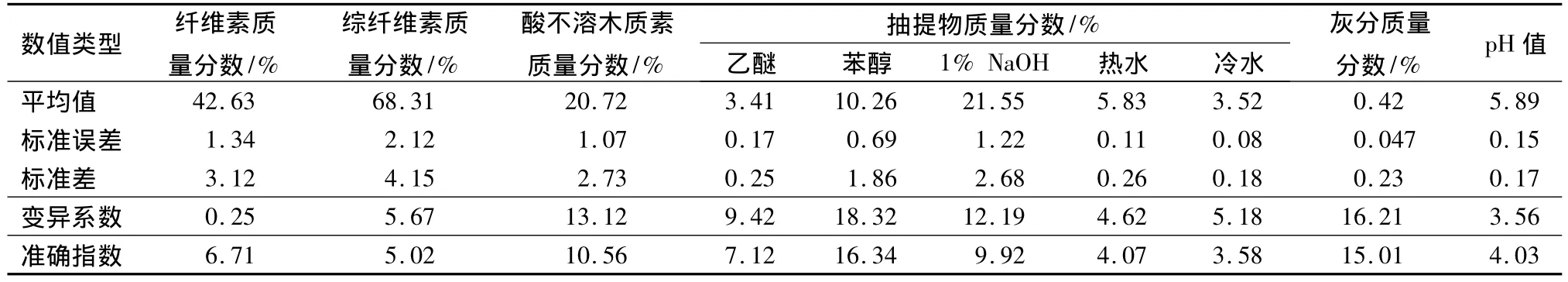
表3 翠柏木材化学性质
3 结论与讨论
在前人对木材解剖性质的大量研究中,主要针对纤维、导管或管胞、薄壁组织和木射线的空间结构做了定性描述和定量分析。衍生出的指标众多,如梁善举等[26]研究米老排(Mytilaria laosensis)纤维和导管的指标就包括长度、宽度、双壁厚、长宽比、壁腔比、腔径比。总体来看,不同生长轮和树高会造成木材纤维、导管或管胞空间结构的变异[26-29]。比如纤维长度、宽度和长宽比自髓心向外随形成层年龄的增大而增大,微纤丝角随形成层年龄递增而递增,而木射线和薄壁组织的变异规律不明显。对木材化学性质研究的指标包括相对结晶度、综纤维素质量分数、木质素质量分数、NaOH抽出物质量分数、糖类质量分数和pH值等。不同树高和生长轮对木材化学性质有一定的影响[30],同时不同种源间的木材化学性质差异显著[31],这方面不同树种所表现出的规律不一。木材纤维素结晶度是衡量木材性质的一项重要指标,是指纤维素的结晶区占纤维素整体的百分比,它反映纤维素聚集时形成结晶的程度。通常随着纤维素结晶度的增加,纤维的抗拉强度等力学性质加强,但是与非结晶区域有关的保水性等物理特性和化学反应性等随之降低[32]。石雷等[25]指出印度黄檀一些木材解剖特征与结晶度存在相关性,并用这些特征与生长轮建立了预测模型。本研究发现不同生长轮对木材的解剖特征以及化学性质有较大影响,这与前人的研究结果相符。此外,进一步把5项木材解剖特征指标(管胞长度、管胞厚度、管胞壁厚、胞壁率和纤丝角)与结晶度进行拟合,得到的模型均为二元三次方程,同类工作目前还未见他人报道。
木材管胞形态的变化受多方面影响,尤其是在树木生长发育的年龄阶段,管胞形态在树木幼龄期变化非常活跃,随着树龄的增大而迅速增加长度,一定年龄后达到稳定。研究表明翠柏木材管胞长度径向变异趋势为20 a前增加且变化显著,之后趋于稳定,且管胞长度从1886 μm增加至3370 μm。依据国际木材解剖学会的规定,翠柏木材管胞属于长纤维,与兴安落叶松(Larix gmelinii)一样是作为制浆造纸等用途的很好的纤维原料[33]。翠柏木材胸径处纤维宽度从髓心到树皮都表现为增加的趋势,与管胞长度径向变异趋势一致,这与不同生长发育时期树木内促进细胞分裂的激素多少有较大关系。翠柏木材胞壁率、微纤丝角和结晶度与其管胞形态径向变异趋势基本一致,均表现为20 a前增加且变化显著,之后趋于稳定。木材结晶度在一定程度上反映了木材纤维的物理和化学性质,翠柏木材结晶度并非与各解剖参数间呈现相关性,与管胞长度、宽度、壁厚和胞壁率呈0.01水平显著正相关,与纤丝角呈0.01水平显著负相关,与组织比量无显著相关。对翠柏木材结晶度与各解剖参数(除组织比量外)进行模型拟合后发现,建立的模型相关系数均在0.9以上。木材的化学性质在一定程度上影响着木材的价值和使用范围,翠柏木材纤维素的质量分数是42.63%,综纤维素的质量分数是68.31%,木质素的质量分数是20.72%;乙醚、苯醇、1%NaOH、热水、冷水的质量分数分别是3.41%、10.26%、21.55%、5.83%和3.52%;灰分的质量分数为0.42%;pH 值为5.89。研究结果认为翠柏是一种较好的用材树种,其测量结果及预测模型可以为合理培育及开发利用翠柏提供基础数据和理论指导。
[1]江泽慧,彭镇华.世界主要树种木材科学特性[M].北京:科学出版社,2001.
[2]刘盛全,江泽慧.人工林的发展和人工林材性与培育及利用关系[J].世界林业研究,1998(4):42-46.
[3]国家环保局,中科院植物所.中国珍稀濒危植物名录:第1册[M].北京:新学出版社,1987.
[4]陈文红,税玉民,王文,等.云南易门翠柏和黄杉的群落调查及保护[J].云南植物研究,2001,23(2):189-200.
[5]宁世江,赵天林,唐润琴,等.木论喀斯特林区翠柏群落学特征的初步研究[J].广西植物,1997,17(4):321-330.
[6]陈子牛.滇中翠柏纯林的生态研究[J].昆明师专学报,1997,12(2):15-22.
[7]谭开湛.保护和发展珍稀树种:翠柏[J].林业调查规划,1982(4):35-36.
[8]张茂钦.翠柏[J].云南林业,1983(3):28.
[9]吕玉华.野生翠柏苗人工容器育苗技术[J].云南林业,2001(3):19.
[10]张学星,何蓉,施莹,等.云南乡土绿化树种对HCl和HF气体的反应[J].西北林学院学报,2006,21(5):47-51.
[11]张学星,何蓉,施莹,等.云南13种乡土绿化树种对SO2、NO2气体反应的研究[J].西部林业科学,2005,34(4):41-46.
[12]刘济明.贵州茂兰喀斯特山地顶部森林群落种子库研究[J].林业科学研究,2000,13(1):44-50.
[13]江泽平,王豁然.柏科分类和分布:亚科、族和属[J].植物分类学报,1997,35(3):236-248.
[14]李玉媛,司马永康,方波,等.云南省国家重点保护野生植物资源的现状与评价[J].云南植物研究,2003,25(2):181-191.
[15]Wang D L,Li Z C,Hao G H,et al.Genetic diversity of Calocedrus macrolepis(Cupressaceae)in southwestern China[J].Biochemical Systematics and Ecology,2004,32:797-807.
[16]中国国家技术监督局.GB 1927-43—91木材物理力学性质试验方法[S].北京:中国标准出版社,1991.
[17]姜笑梅.人工林与天然林杉木幼龄材和成熟材解剖性质差异及其在径向上变异的比较研究[J].世界林业研究,1994,7(专集):1-13.
[18]Segal L,Creely J J,Martin A E,et al.An empirical method for estimating the degree of crystallinity of native cellulose using the X-ray diffract-meter[J].Textile Research Journal,1959,29:786-794.
[19]中国标准出版社.中国林业标准汇编:木材与木制品卷[M].北京:中国标准出版社,1998.
[20]北京造纸研究所.造纸工业化学分析[M].北京:轻工业出版社,1975.
[21]Schimleck L R,Evans R.Estimation of wood stiffness of with cement cores by near withdrawer spectroscopy:the development and application of calibrations based on selected cores[J].IAWAJ,2002,23(3):217-224.
[22]Panshin A J,De Zeeuw C.Textbook of wood technology[M].New York:McGraw-Hill Book Company,1980.
[23]Zobel B J,Bitumen J P.Wood variation:its causes and control[M].Berlin Heidelberg:Springer-Verlag,1989.
[24]Lee C L.Crystallinity of wood cellulose fibers studies by X-ray methods[J].Forest Products Journal,1961(11):108-112.
[25]石雷,孙庆丰,邓疆.人工幼龄印度黄檀木材解剖性质和结晶度的径向变异及预测模型[J].林业科学研究,2009,22(4):553-558.
[26]梁善庆,罗建举.人工林米老排木材解剖性质及其变异性研究[J].北京林业大学学报,2007,29(3):142-148.
[27]虞华强,费本华,赵荣军,等.中山杉和落羽杉木材解剖性质研究[J].林业科学研究,2007,20(2):213-217.
[28]潘彪,徐朝阳,王章荣.杂交鹅掌楸木材解剖性质及其径向变异规律[J].南京林业大学学报:自然科学版,2005,29(1):79-82.
[29]邱帖轶,常德龙,唐君畏.秃叶黄皮树的木材解剖性质[J].东北林业大学学报,2006,34(3):36-38.
[30]周亮,刘盛全,高慧,等.欧美杨107杨木材化学组成径向变异[J].东北林业大学学报,2010,38(12):10-14.
[31]汪贵斌,曹福亮,柳学军,等.不同落羽杉种源木材化学性质的变异[J].南京林业大学学报:自然科学版,2009,33(6):15-19.
[32]杨淑敏,江泽慧,任海青,等.利用X射线衍射法测定竹材纤维素结晶度[J].东北林业大学学报,2010,38(8):75-77.
[33]白默飞.人工林兴安落叶松生长应力与木材性质关系的研究[D].合肥:安徽农业大学,2009.
猜你喜欢
军事文摘(2021年16期)2021-11-05
中学生数理化·高一版(2021年3期)2021-06-09
数学物理学报(2021年1期)2021-03-29
少儿科学周刊·少年版(2021年20期)2021-01-17
建材发展导向(2020年16期)2020-09-25
重型机械(2020年3期)2020-08-24
数学年刊A辑(中文版)(2019年3期)2019-10-08
核技术(2016年4期)2016-08-22
江苏农业科学(2015年1期)2015-04-17
中国质量与标准导报(2014年5期)2014-02-28
