
甘菊种子萌发特性及自然条件下生长发育规律1)
2012-09-18 01:27付建新戴思兰
东北林业大学学报 2012年4期
丁 焱 付建新 戴思兰
(国家花卉工程技术研究中心(北京林业大学园林学院),北京,100083)
高等植物的生长发育涉及种子萌发、营养生长、开花、受精、胚胎发育、子代种子形成等一系列发育阶段[1]。种子萌发是植物生长发育的起始,在整个生命活动周期中非常关键。影响种子萌发的环境因子较多,其中温度[2-3]、光照和土壤湿度是影响种子萌发最重要的3个环境因子。植物种子由于自身存在的原初休眠和二次休眠以及为了躲避不利的生存环境,往往存在强迫休眠现象[4]。如果栽培和育种工作者不了解种子的休眠特性,找不到合适的方法打破种子休眠,就会给农业、林业生产带来极大的困难[5]。生长是指植物体积和质量不可逆的增加,常指营养生长;发育则强调植物的质变,常指生殖生长。环境因子,如温度、光照、水分、养分、土壤及根际环境和空气成分,对植物生长发育均有不同程度的影响[6]。只有掌握园林植物的生长发育过程才能正确地使用园林植物,发挥它们在园林中的作用。
甘菊[Chrysanthemum lavandulifolium(Fisch.ex Trautv.)Makino]又名岩香菊,为多年生草本植物,高0.3~1.5m,有地下匍匐茎。秋季黄花繁密,耐粗放管理,已渐渐被引入到园林绿化中[7]。甘菊是菊花起源的重要亲本之一,其为二倍体,遗传背景比较简单,开花期与菊花一致,为典型的短日照成花植物,是菊属植物分子生物学研究的理想模式植物[8-12]。目前已有很多关于菊科植物栽培和遗传转化方面的研究报道[13-18],但对甘菊生物学特性的研究还较少。对甘菊种子的萌发特性和自然条件下生长规律进行研究,可以为将甘菊开发成菊科植物研究的模式材料奠定良好基础,并为甘菊再生及转化体系、分子育种等方面的研究工作提供材料及依据。
1 试验地概况
试验地设在北京林业大学苗圃,位于北纬39°54',东经116°24',海拔50 m。属暖温带半湿润半干旱大陆性季风气候,冬季寒冷,夏季炎热。年平均气温11.8℃,最热月为7月,月平均气温26.1℃;最冷月为1月,月平均气温为-4.7℃。年无霜冻期190~200 d。年平均降水量639.8 mm,且多集中在夏季。
2 材料与方法
试验材料:甘菊种子于2006年11月25日及2007年11月23日采自北京师范大学、北京大学、北京林业大学及北海公园等地。
甘菊种子萌发实验及发芽率的测定:甘菊种子采收后,于28℃恒温烘箱中烘干,去除杂质后常温避光保存在牛皮纸袋中,空气相对湿度为20% ~30%。次年春季分别在培养皿、MS培养基中和盆栽园土中播种,观察其发芽率。筛选出甘菊种子萌发最佳培养基质后,分别将甘菊种子于当年秋季、次年春季、次年秋季和第三年春季4个时间段在甘菊种子萌发最佳培养基质中播种,观察采收后时间长短对甘菊种子萌发率的影响。种子萌发的环境条件设置为温度24℃,光照强度2400~2600 lx,光周期16 h/8 h。胚根长度达到0.5 cm作为发芽标准,20 d后统计发芽率。每处理均设3次重复,每个重复30粒种子。
甘菊周年生长状况调查:在北京林业大学苗圃地里正常生长的人工播种苗中随机选取1年生幼苗10株,于2007年5月至10月定期测量甘菊植株高度、茎干粗度、冠幅变化、叶片数量和分枝数,并进行统计分析。
甘菊年生长温度及光周期观测:采用温度计测量甘菊年生长期间温度并记录分析。根据国旗升降时间记录自然日照长度,观察并记录试验地甘菊营养生长期、现蕾期和开花期,分析甘菊不同生长发育时期对应的日照长度和温度。
3 结果与分析
3.1 甘菊种子萌发率
甘菊种子为瘦果,长1.2~1.5 mm,长椭圆形,有肋,每粒质量约0.139 mg。不同培养基质对甘菊种子萌发率有显著性影响,其中甘菊种子在MS培养基中萌发率最高,达到93%,而在园土中撒播的种子萌发率仅有80%(表1)。采收后立即播种和春季播种两种处理条件对甘菊种子萌发率没有显著性影响,种子萌发率均保持在90%左右,而第二年秋季播种,萌发率迅速下降,种子存放2 a后,萌发率下降到30%以下(表2)。
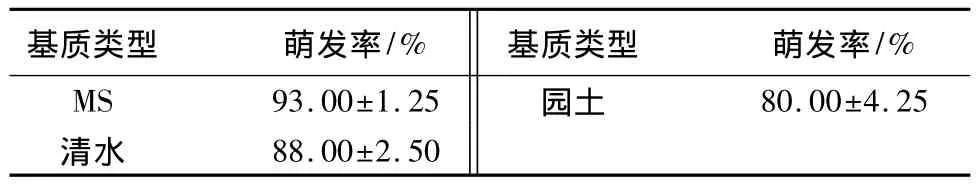
表1 不同栽培基质对甘菊种子萌发率的影响
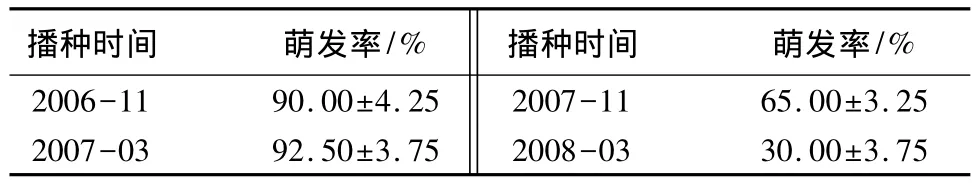
表2 不同储藏时间对甘菊种子萌发率的影响
3.2 自然条件下甘菊年生长状况
通过田间观测可以看到,5—8月份是甘菊植株快速高生长时期,株高变化很快,而9月份高生长增长开始变慢,至10月份株高增长更加不明显(表3);此期间,甘菊茎粗增长最为明显,从1 mm迅速变为1.75 mm,9月份甘菊茎粗达到生长最大值(2.92 mm),9月中旬—10月甘菊茎粗增长缓慢,1个月时间仅增加0.06 mm(表3);5—8月份甘菊植株叶片数量迅速增加,9月份叶片数量变化不明显,从10月开始,甘菊叶片数开始下降,这主要是由于甘菊进入衰老阶段,老叶逐渐脱落的缘故(表3)。5—6月份甘菊冠幅变化最大,9月份以后则变化不明显,10月份以后平均冠幅也有轻微下降趋势(表3)。5月中旬到6月份甘菊分枝数很少,6月中旬至8月中旬甘菊分枝数迅速增加,9月份甘菊分枝数增长缓慢,10月中旬甘菊分枝数与9月份基本一致,不再变化(表3)。
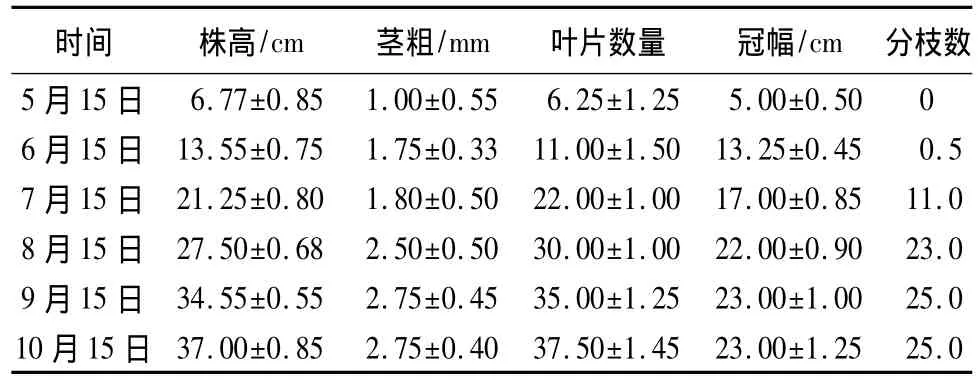
表3 自然条件下甘菊年生长状况
3.3 自然温度及光照条件下甘菊年生长发育规律
自然条件下,甘菊种子于3月中下旬开始萌动,种子萌动后甘菊叶片迅速生长,5月初甘菊进入快速生长阶段。经过4个月左右的营养生长,9月中旬开始现蕾,9月下旬至10月底为甘菊盛花期并产生萌蘖。甘菊植株中部以上多花枝或伞房状花序分枝,头状花序在茎顶端形成复伞房花序,舌状花黄色,靠上部花序先行开放,然后逐次向下和两旁开放。单花花期一般5 d,群体花期达30 d以上。l1月初至12月初结实,同时地上部分枯黄倒伏,直至完全枯萎,甘菊从此进入休眠期(表4)。
4 结论与讨论
不同栽培基质对甘菊种子萌发率影响不大,其中以MS培养基中播种的甘菊种子萌发率最高,但其操作较为繁琐,如需大量种植,可以选取培养皿培养萌发后,将幼苗用镊子夹入基质中或在园土中直接撒播进行种植。甘菊种子没有休眠期,选择采收后立即播种或第2年早春播种,发芽率均较高,但储存2 a及以上的种子,生活力明显下降,萌发率很低。对甘菊种子的储存技术还需要探讨,目前的研究只能采用每年收集的新鲜种子,或以盆栽植株保存材料。
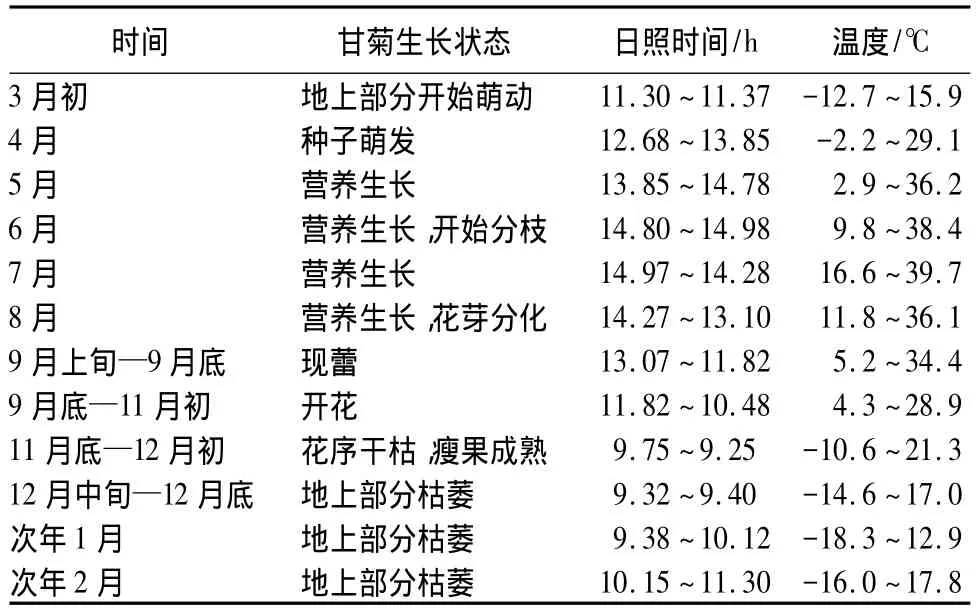
表4 甘菊年生长状态调查结果
甘菊在自然条件下,5月份到8月中旬叶片快速生长,到10月份,叶片数量微微下降,这是由于甘菊到了生殖期,不再进行营养生长,叶片不再增加,而下端叶片干枯甚至部分脱落造成的。5月份至6月份甘菊冠幅变化最大,这可能是由于此时甘菊植株已经脱离幼态,开始迅速生长,叶片明显变大,致使冠幅变化明显;而9月份以后甘菊冠幅变化不明显,这是由于甘菊植株不再进行营养生长,而部分叶片开始萎蔫干枯的缘故。甘菊分枝数的变化曲线呈现出慢-快-慢的变化趋势,5月份至6月中旬变化不明显,6月中旬以后分枝数迅速增加,直到9月,之后分枝增加速率迅速下降,10月份由于甘菊已经进入盛花期,分枝数保持不变。苗期甘菊植株在6月份以后开始迅速进行高生长,一直持续到8月后,因为转为生殖生长,所以生长量变慢。植株所呈现的生长规律与生长环境密切相关。北京地区地处中纬地带,气候是明显的暖温带、半湿润大陆性季风气候。冬、夏时间长,而春、秋时间短。春季天气较冷,多风,雨水少,天气干旱,不利于幼苗发育,导致植株生长缓慢,而夏季合适的温度和充沛的降雨利于植株幼苗旺盛生长。秋季植株基本停止营养生长,开花结实后开始发脚芽,母株不再生长,直至发黄干枯。
高等植物的生长发育过程主要包括营养生长和生殖生长两个阶段。自然条件下,5—8月份是甘菊营养生长的快速生长期,在此期间日照时间为13 h以上,平均温度为23.85℃;9—10月份甘菊进入生殖生长阶段,当日照长度缩短至13 h时甘菊开始现蕾,当日照长度缩短至12 h左右时甘菊开始开花,生殖生长期间平均温度为16.55℃。因此,在人工环境中可以通过温度和光照时间调控甘菊生长发育过程。通过本研究可以推测甘菊营养生长适宜的光周期条件为14 h光照/10 h黑暗,生殖生长适宜的光周期条件为12 h光照/12 h黑暗,此结果还需更多试验进一步验证。
模式植物是指生物的一个物种,它在人们研究生命现象的过程中长期反复地被作为研究材料,并且从这个物种的研究中得出的许多生命活动规律代表许多物种的共同规律[19]。通常模式植物需具备以下条件:易于培养,成本低廉,繁殖周期短,短时间可以产生大量后代,方便取得种内遗传变异体等。目前常用的模式植物主要有拟南芥、水稻、玉米、葡萄等。甘菊是菊科菊属甘菊系多年生二倍体草本植物,是菊花起源的重要亲本之一,遗传背景简单[20]。甘菊是北方常见的野花,抗旱抗寒能力强,可以自播繁衍,材料易于获得[21]。对其基本生物学特性进行研究可以为菊科植物的研究提供一定的指导作用,因此,甘菊有望成为菊科植物遗传育种研究的模式植物。
通过对甘菊种子萌发特性和自然条件下生长规律进行研究,获得了甘菊自然生长条件下年生长发育规律,为今后对甘菊开展开花期、抗逆性等相关基因的表达及功能验证的研究,为甘菊成为菊科植物的模式植物奠定了良好的基础。
[1]付建新,王翊,戴思兰.高等植物CO基因研究进展[J].分子植物育种,2010,8(5):1008-1016.
[2]张鹤山,陈明新,田宏,等.高温胁迫下白三叶种子萌发特性及耐热性研究[J].种子,2010,29(8):1-5.
[3]蒋小军,文向多.温度对蓖麻种子萌发的影响[J].种子,2008,27(5):67-69.
[4]孙海莲,阿拉塔,王海明.华北驼绒藜种子萌发特性的研究[J].内蒙古草业,2010,22(3):47-50.
[5]周霞,张尧.种子休眠生理研究进展综述[J].安徽农学通报,2010,16(14):51-53.
[6]刘燕.园林花卉学[M].1版.北京:中国林业出版社,2003.
[7]曹华雯,刘振林,夏新莉,等.甘菊DlBADH1基因启动子DBP12表达特性研究[J].分子植物育种,2007,5(6):758-764.
[8]刘振林,尹伟伦,戴思兰.新的BADH同源基因:甘菊BADH基因[J].分子植物育种,2005,3(4):591-593.
[9]Ma Y P,Fang X H,Chen F,et al.DFL,a FLORICAULA/LEAFY homologue gene from Dendranthema lavandulifolium is expressed both in the vegetative and reproductive tissues[J].Plant Cell Rep,2008,27:647-654.
[10]刘振林,冯慧,戴思兰.北京及周边地区菊科野生花卉资源调查与分析[C]//张启翔.中国观赏园艺研究进展.北京:中国林业出版社,2005:69-73.
[11]刘慎谔.东北植物检索表[M].北京:科学出版社,1995:20.
[12]丁焱,戴思兰.甘菊再生体系的建立[C]//张启翔.中国观赏园艺研究进展.北京:中国林业出版社,2005:219-221.
[13]洪波,史春凤,张晓娇,等.菊花观赏性状和农艺性状基因工程改良研究进展[J].中国农业科学,2009,42(4):1348-1358.
[14]王春彦,高年春,张效平,等.非洲菊组织培养工厂化育苗关键技术研究[J].南京农专学报,2003,19(2):32-37,42.
[15]洪波,张常青,李邱华,等.根癌农杆菌介导的转录因子DREB1A基因在地被菊花中的遗传转化[J].农业生物技术学报,2005,13(3):304-309.
[16]蒋细旺,包满珠,吴家和,等.农杆菌介导Cry1Ac基因转化菊花[J].园艺学报,2005,32(1):65-69.
[17]洪波,仝征,李邱华,等.地被菊花Fall Color体细胞胚途径再生、遗传转化及转基因植株的抗寒性检测[J].中国农业科学,2006,39(7):1443-1450.
[18]郑丽,李名扬,晁岳恩,等.根癌农杆菌介导ipt基因对切花菊的遗传转化[J].农业生物技术学报,2005,13(1):26-31.
[19]曹仪植.拟南芥[M].1版.北京:高等教育出版社,2004:3.
[20]戴思兰.中国栽培菊花起源的综合研究[D].北京:北京林业大学,1994:19.
[21]马月萍.甘菊LEAFY同源基因的克隆与表达分析[D].北京:北京林业大学,2005:17.
猜你喜欢
热带林业(2022年4期)2023-01-24
矿山安全信息(2022年22期)2022-11-24
中国农业科学(2022年12期)2022-06-28
山西农业科学(2020年5期)2020-05-19
江西农业(2020年11期)2020-02-12
中国新闻周刊(2019年46期)2019-12-20
作文周刊·小学三年级版(2019年48期)2019-02-02
天津农业科学(2017年5期)2017-05-15
长江蔬菜·学术版(2014年4期)2014-05-09
植物营养与肥料学报(2011年4期)2011-10-26
