
厚荚相思家系抗寒性变异与选择1)
2012-09-18 01:27黄银珊杜加加
东北林业大学学报 2012年4期
梁 机 李 婷 黄银珊 杜加加
(国家林业局中南速生材繁育重点实验室(广西大学),南宁,530004)
樊东函 陈乃明
(广西钦州市林科所)
厚荚相思(Acacia crassicarpa)为含羞草科金合欢属的热带乔木树种。原产澳大利亚昆士兰东北沿海及内地,巴布亚新几内亚西部省及印度尼西亚伊里安岛也有分布[1]。该树种具有生长迅速,对立地要求不严的特性,其根系发达,且具根瘤能固氮改土功能,可作为防治林地因过渡连栽而引起地力衰退的理想树种。我国于20世纪70年代末开始引种栽培,近年来在我国南方地区如福建、广东、广西等地推广种植,已逐渐成为在纸浆材、建筑材、家具材、薪炭材、水土保持、荒山绿化以及生态恢复和生态重建等方面得到广泛应用的又一重要的造林树种[2-3]。然而相思类树种不耐寒,限制了其进一步向北推广种植,低温冻害是厚荚相思扩大栽培与速生丰产的主要限制因子[4-5]。因此,选择耐寒性强的品种成为目前降低栽种风险的主要措施之一。2008年初广西遭受了历史罕见的雨雪冰冻及连续低温寒害天气袭击,使营林区内的相思树林分遭受不同程度的寒害,但是灾后在我们所营建的厚荚相思种子园及其子代测定试验林中发现尽管大部分植株受寒害作用甚至死亡,仍有部分家系的植株完全未受害,表现出高度的抗寒性。幼林阶段的林木因其枝、叶比较嫩弱,使幼树的适应性和抗逆性往往差于成年植株,所以对幼树的抗寒能力选择与研究将更具有实用价值。为探索厚荚相思不同家系在抗寒能力上的差异及抗寒性的遗传变异规律,本文通过对两个试验点的厚荚相思子代测定林进行田间抗寒性调查测定,研究揭示其幼林不同家系间的抗寒性差异,以期为选育抗寒能力较强的厚荚相思品系,提高抵御自然灾害的能力,扩大其栽培推广范围,实现适地适树营林提供理论依据。
1 试验地概况
试验点1设在广西高峰林场大塘分场,位于南宁市的东北部,北纬 22°58',东经 108°32',属于亚热带季风气候区。年平均气温21.4℃,极端最高气温40.4℃,极端最低气温-2.1℃。霜期短,年平均有霜期仅4.3 d,年降水量为1300 mm。地貌主要为丘陵和山丘构成,试验地坡度较陡达25°左右,坡向为北坡。子代试验林采用完全随机区组设计,4次重复,5株小区,株行距为2 m×2 m。
试验点2设在钦州市林科所,位于北纬21°57',东经108°33',属亚热带海洋性季风气候区,具有热带向亚热带过渡的特点,年平均气温21.5℃,年总积温7850~8030℃,冬无严寒,夏无酷暑,年均降水量1750 mm。地形地势开阔平坦,低丘和台地地貌,土壤为砖红壤。试验点地块完整,坡度小于5°,立地条件基本一致。子代试验林采用完全随机区组设计,3次重复,15株小区,株行距为3 m×2 m。
2 材料与方法
1.1 试验材料
2006年6月从广西钦廉林场125个家系构成的厚荚相思实生种子园内,按表型选择生长及干形性状表现最优良的38株母树(家系)单系采种育苗,以普通生产混合种作对照(CK),2007年3月分别在广西高峰林场大塘分场和钦州市林科所两个地点营造子代测定试验林。
1.2 观测标准与数据分析
1.2.1 不同寒害症状及等级标准
试验参照中华人民共和国国家标准《林木引种》(GB/T 14175—93),根据厚荚相思幼林受冻后出现的症状制订了相应的等级标准,如表1。

表1 寒害分级标准
1.2.2 调查测定方法
2008年1月12日至2月18日,广西连续遭受低温雨雪冰冻天气袭击,发生这次灾害性天气期间,降温强度大、影响范围广、持续时间长。灾害天气过后于2008年3月,对两个试验点遭受严重寒害的1年生厚荚相思子代测定试验林进行全林检尺调查,使用测高杆、电子游标卡尺按小区(家系)进行树高、胸径和冠幅测定,并按照表1厚荚相思寒害等级划分的标准逐株观察记录寒害等级。
1.2.3 寒害程度的计算及数据统计
按家系统计不同受冻级别的株数,采用公式¯a=∑a·n/N计算各家系的小区平均寒害级值;按公式δ=(∑a·n)(amax·N)×100%计算不同家系的寒害指数,以分析不同家系间受寒害相对轻重程度;用公式L=∑n/N×100%计算寒害率。式中:¯a表示平均寒害级别;δ表示寒害指数;a表示受寒害级别(0级为0,Ⅰ级为1,Ⅱ级为2,余此类推);n表示某受寒害级出现的株数;amax表示寒害级值标准中最高级别的级值;N表示调查总株,L表示寒害率。
以小区单株调查值为单位,计算各性状指标在家系间的统计值,统计分析模型为:yijk=μ+bi+fj+bfij+eijk,式中:yijk为第i个区组第j个家系的第k个观测值;μ观测值总体均数;bi为重复效应;fj表示家系效应;bfij表示家系与重复的互作效应;eijk为剩余项。对各性状的家系间变异采用双因素交叉分组的线性统计模型分析,各变异来源的方差分量用混合模型分析,其中区组为固定项,家系为随机项。方差分析及期望均方组成参见参考文献[6]。
遗传力及遗传增益采用以下公式估算[6]:

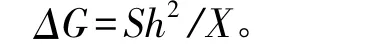
式中:ΔG为遗传增益,S为选择差,h2为家系遗传力,X为群体平均值。
寒害级值、寒害指数在进行方差分析前分别应用平方根、反正弦平方根转换以使数值方差齐性[7],采用DPS7.05统计软件进行试验数据的处理分析。
3 结果与分析
3.1 不同家系的受害程度及抗寒性表现
田间测定是评价林木越冬抗寒性最直观可靠的方法。对遭受了2008年初寒害天气过程的两个试验点厚荚相思子代林进行全林调查测定,结果见表2、表3。
由调查结果可见,两个地点的厚荚相思幼林受寒害程度不同,大塘试点的林分植株普遍受害严重,各家系平均寒害级值在2.67~4.00,寒害指数为66.7% ~100%,平均达92.4%。而钦州试验点的幼林受害相对较轻,寒流过后基本恢复生长,各家系平均寒害级别值为0.60~1.93,寒害指数在15.0% ~48.3%,平均寒害指数为26.0%。
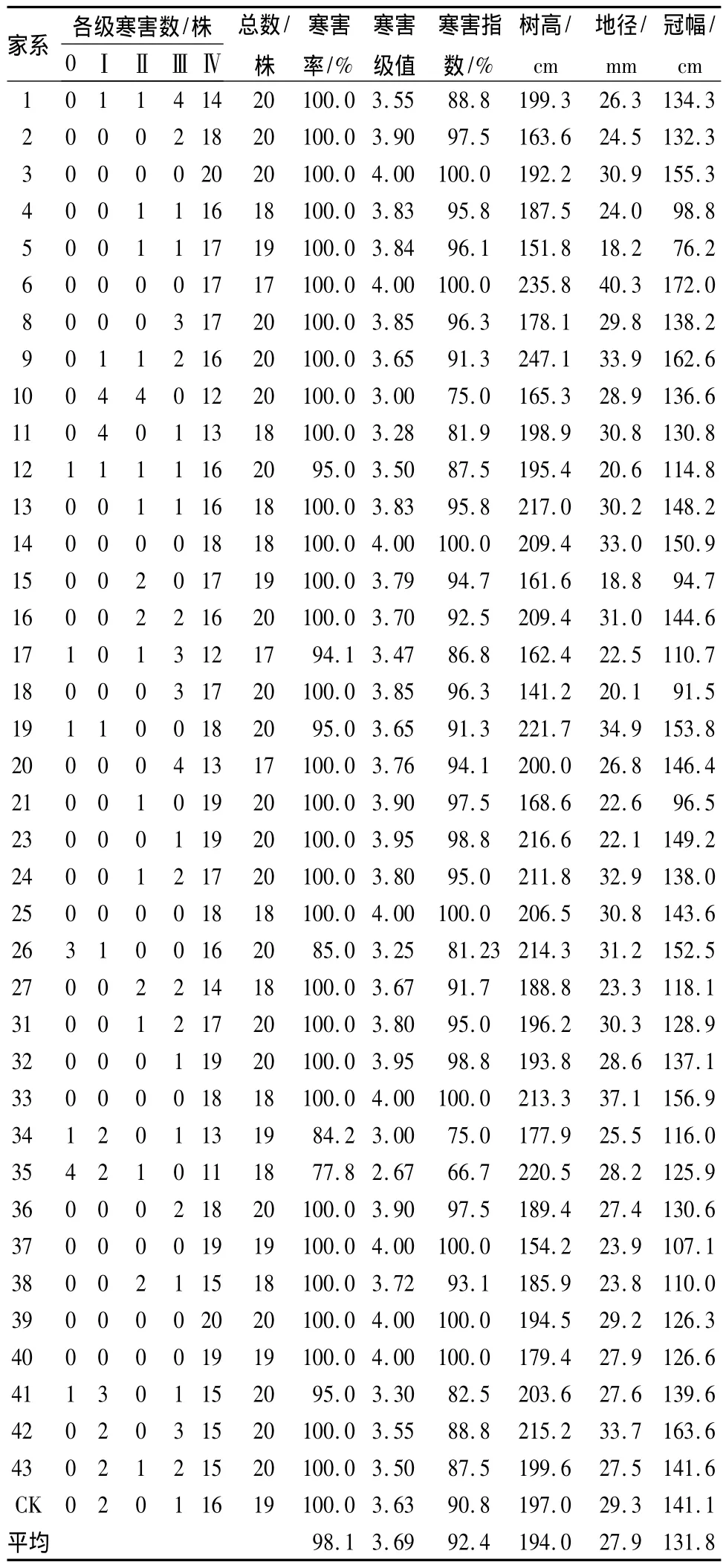
表2 大塘试点不同家系厚荚相思抗寒性比较
在大塘试点参试的39个家系中,受寒害级别Ⅲ级以上寒害程度的家系有37个,占94.9%,受害植株数达707株,占总调查株数的94.6%,其中3、6、14、25、33、37、39、40 号等 8 个家系的寒害级别值达到了4.0,寒害指数达100%,这些家系受害严重,植株全部被冻死亡,表明这些家系对寒害表现出高度敏感性,抗寒性最差。田间调查中发现尽管大部分植株均受到不同程度的寒害,但仍有少量家系植株依然翠绿挺拔,完全未受冷害,表现出高度的抗寒性。寒害指数是评价植物种(品种)间抗寒力水平的重要指标,在相同的寒害环境下,寒害指数越小,抗寒性越强,反之则植物对寒害越敏感。本研究的大塘试验点中,若按寒害指数及寒害级值大小进行排序,则可初步选出抗寒性较强的前10个家系,受害程度大小的排序为35<10<34<26<11<41<17<12<43<1,对寒害作用表现敏感、抗寒能力最差的10个家系分别是 40、39、37、33、25、14、6、3、33、23。

表3 钦州试点不同家系厚荚相思抗寒性比较
在钦州试验点,1年生厚荚相思子代测定林分整体上长势比大塘试点的好,平均树高达到210.5 cm、平均地径为36.7 mm、平均冠幅达到174.1 cm,分别比大塘点高出8.5%、31.5%和32.1%。试验林中没有一个家系的平均寒害级别值达到Ⅲ级以上寒害程度,平均寒害指数比大塘点低66.4%,在参与调查的1664株植株中,受Ⅲ级和Ⅳ级严重寒害的株数仅出现77株,占总调查株数的4.6%。根据寒害指数和寒害级值的大小,该试验点参试家系中受寒害轻、表现抗寒能力最强的前10个家系为10<34<35<41<9<43<26<19<14<11,而对寒害作用表现敏感、抗寒能力较差的10个家系分别是40、25、21、39、23、32、37、33、13、36。
3.2 不同家系抗寒水平的差异性
为分析厚荚相思子代各家系抗寒水平的差异性,以家系的小区单株寒害级值为样本值对其抗寒性状的家系间变异进行双因素交叉分组的线性统计模型分析,方差分析结果(表4)表明,无论是在大塘试点还是钦州试验点,参试的39个家系子代其抗寒性在家系间均达到极显著水平(p<0.01),说明来自不同家系的厚荚相思材料,在抗寒水平上存在明显的差异,这为厚荚相思家系子代进一步在抗寒力性状上的选择提供了可行性。
从方差分量可知,两试验点内寒害级值的遗传变异主要来源是家系与区组的交互作用以及小区内的变异,均占到总方差的75%以上,在钦州点的家系方差分量中,剩余项>互作>家系,可见家系内单株间的变异分量比重较大,表明在家系内进行个体选择有较大潜力。两地点寒害级值的家系方差分量分别为9.922%和15.447%,进一步估算寒害级值的家系遗传力和单株遗传力分别为0.863、0.941和0.414、0.683,显示家系间抗寒能力呈高强度遗传,表明抗寒性状在较大程度上受遗传效应的控制,家系遗传力均高于单株遗传力,通过在家系层次上的选择比基于单株的选择更能获取较大的遗传增益。

表4 家系间寒害级值方差分析、方差分量及遗传参数的估算
3.3 抗寒性的地点间及与生长性状相关性
对两个试验点间幼林的寒害程度及试验林内不同家系的抗寒性与树高、胸径、冠幅等生长因子的相关性采用Pearson Correlation相关性检验(表5),结果显示不同地点间各家系厚荚相思幼林的寒害率和寒害指数均呈极显著正相关,表明了各家系的抗寒性状在不同地点表现较稳定一致,通过对比两试验点内各家系的抗寒表现,有望比较可靠地选出具有强抗寒能力的共同家系。同一地点内寒害率和寒害指数与树高、地径和冠幅的生长没有明显的相关关系,但树高、地径和冠幅各生长性状间则呈现极显著的正相关。
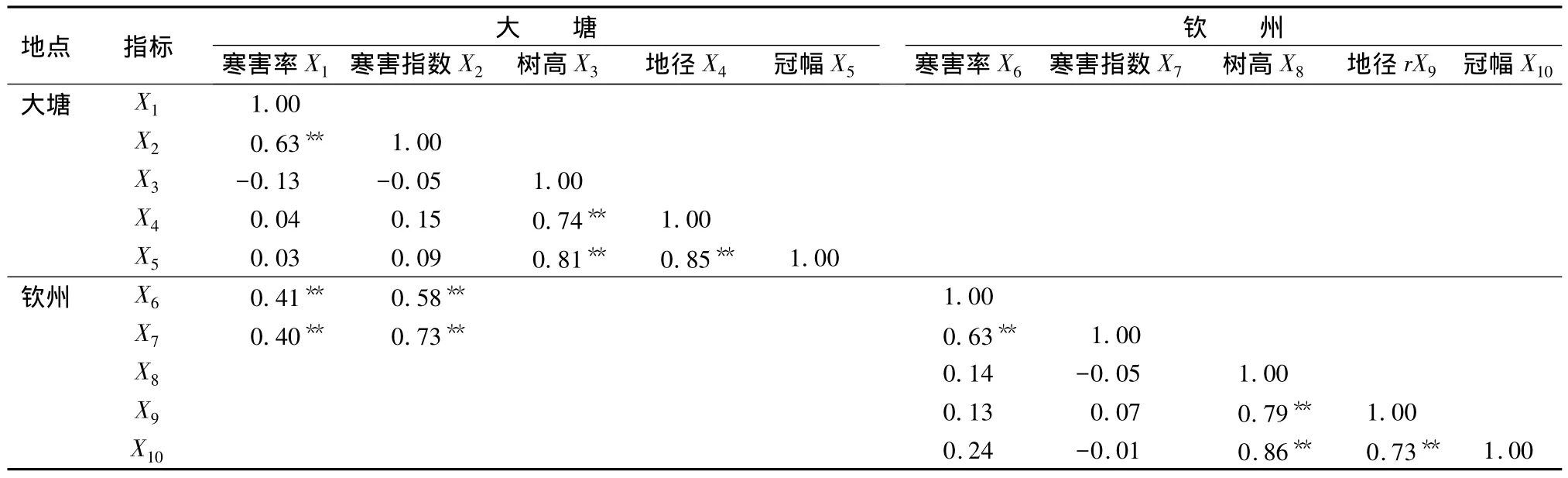
表5 厚荚相思家系抗寒性与生长因子相关性分析
3.4 抗寒家系选择及遗传增益的估算
若按寒害指数及寒害级值大小进行排序分别选出2试验点最耐寒的前10个家系(表6),则大塘点和钦州点选中家系的寒害指数小于群体平均值分别为3.9% ~27.8%、15.8% ~42.3%,选中家系的寒害指数平均值分别比对照(CK)低11.6%和11.7%,抗寒性状寒害级值的平均遗传增益分别可达10.23%和 21.89%。
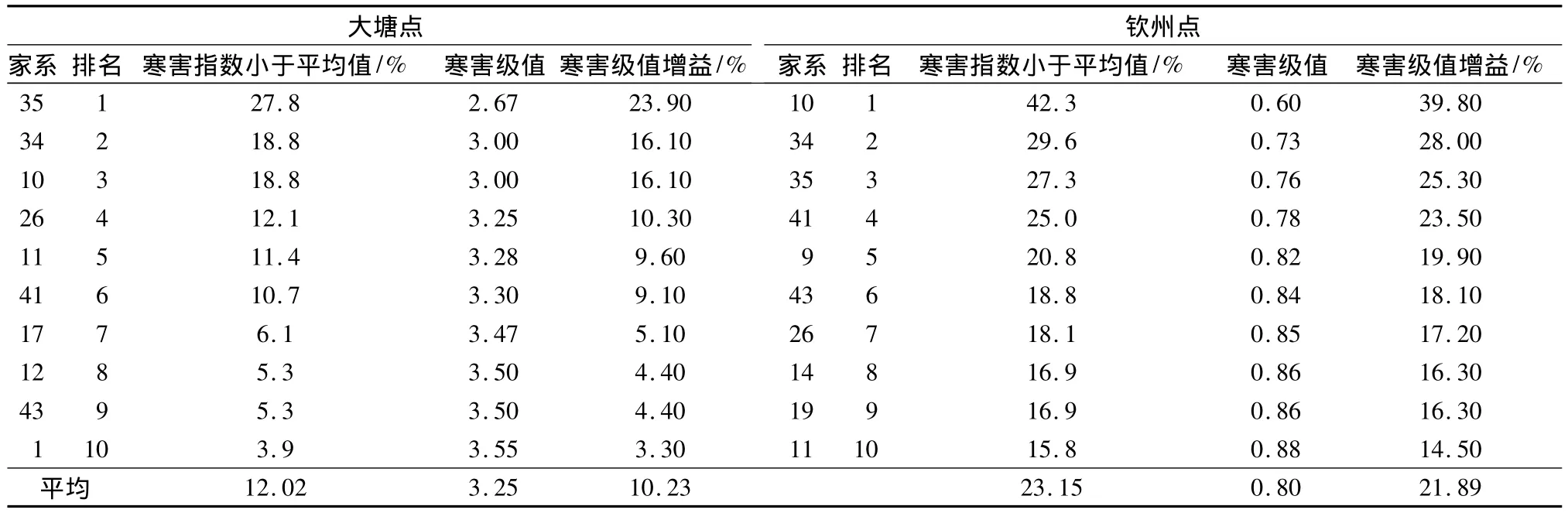
表6 耐寒性排序前10名家系及其遗传增益
4 结论与讨论
4.1 两个试验地植株寒害差异与主要致害气象因子
试验点不同的地理纬度及地形地貌是导致两地植株寒害差异的主要因素。设立在两个不同地点的1年生厚荚相思子代测定林在2008年初经历了连续低温雨雪冰冻天气过程后,所有参试家系均受到寒害影响,但不同地点试验林分的受害程度表现不一,位于南宁市的大塘试验点林木受害最严重,各家系平均寒害级别值在2.67~4.00,平均寒害指数达92.34%。而设在钦州市林科所试验点的幼林受害相对较轻,各家系平均寒害级别值为0.60~1.93,平均寒害指数为26.0%。厚荚相思为热带树种[1],由于其长期适应热带亚热带的气候特点,对低温的抵抗能力比较差[4-5],当遭受广西50 a一遇的强寒流袭击时均表现出寒害反应。尽管南宁的大塘试点属于亚热带季风气候区,北回归线以南,冬季通常在10℃以上,气温温和,正常年份较少发生寒害,但该试验点内可达-2.1℃的极端低温及较陡峭的山丘地形常出现的强辐射降温往往是导致林木主要寒害的气象因子,尤其是在2008年1月12日—2月18日,出现了以2~7℃的低温为主,低于10℃的低温天气长达37 d,持续低温是造成大塘试点的厚荚相思受害严重的主要原因,此外在持续低温过程后又有一个气温急速回升又骤降的过程,由4~6℃迅速回升至25~30℃,气温的大幅度波动更加剧了植株的寒害程度,因而导致该试验点受寒害程度Ⅲ级以上的家系占94.9%,受害植株数总调查株数的94.6%,且有8个家系全部被冻死亡。而钦州市地处亚热带海洋性季风气候区,具有热带向亚热带过渡的特点,位于北纬22°58',其地理位置相对于南宁略靠南,纬度低1°,虽然也遭受寒害,但持续时间和降温幅度没有南宁长,平均气温低于10℃的时间为9 d,因此设在钦州试验点的厚荚相思子代林受害程度要相对轻于大塘试点。
4.2 家系间和家系内抗寒性差异的遗传效应及选择利用
不同试验点中厚荚相思子代林家系间和家系内两个不同变异层次上的抗寒水平均存在极显著差异,说明可以从参试家系中选择得到耐寒力强的优质种质。寒害级值的家系遗传力达0.863~0.941,表明厚荚相思家系间抗寒性状的差异是遗传因素引起的,这为从家系中选择高抗寒性优良家系提供了基础。遗传变异分析结果显示,家系内单株间的变异分量占总方差的比重较大,是家系间变异量的2~4倍,说明家系内部单株的抗寒性差异十分明显,这意味着在家系内进行个体选择有较大潜力。因此,在选择过程中可结合改良性状的遗传力,在以多个性状综合评定值的高低来选择出最优良家系的基础上,通过家系—单株双层次联合选择,进而获得更好的选择响应。近年来许多学者通过不同的抗寒性测定方法在树木抗寒机理、抗寒性鉴定、抗寒资源评价及抗寒性遗传育种等领域进行了大量的研究,试验证明林木在种源间[8-10]、家系间[11-12]、无性系间[13]、品种(类型)甚至单株个体间[14]存在显著的抗寒性差异,可见植物抗寒力性状的遗传变异现象广泛存在于林木群体中。据此,不少研究者根据田间鉴定、电导率测定、生理生化指标测定等比较成熟的抗寒性鉴定技术筛选出了许多优良的高度抗寒林木品种和果树砧木资源[15]。本研究结果显示厚荚相思子代各家系间抗寒水平差异显著,不同家系寒害指数变幅宽,这为厚荚相思家系子代进一步在抗寒力性状上的选择和改良提供了可行性,经历了此次特大寒流袭击后仍然保存完好的参试家系与单株,也为开展厚荚相思耐寒选育改良提供了难得的机遇及基础育种资源。
4.3 抗寒性与生长指标的相关性
厚荚相思子代林各家系的寒害率和寒害指数两地点间的相关系数分别为0.41和0.73,达到极显著水平的正相关,表明耐寒能力具有较高的遗传稳定性。从子代林各家系的寒害指数和平均寒害级别值排序来看,两个试验点各家系的抗寒性排名次序虽不完全相同一致,但两地点共同对寒害表现高度敏感的前10个家系和表现高度抗寒性的前10个家系基本包括在内。子代林各家系的抗寒指标与生长性状的相关系数都很小(0.01~0.24),未达显著水平。有关对林木抗寒性与生长性状相关分析研究的结果报道不一,邹小兴等[13]的研究表明巨桉种源和家系冻害指数与树高、胸径和材积各性状之间为极显著负相关,王家源等[7]在不同产地苦楝苗木抗寒性差异的试验中观察到同一产地内苦楝苗木受冻害程度与地径生长达极显著的负相关水平,而不同产地苦楝苗木受冻害程度则与其平均地径呈极显著正相关。本研究发现同一地点内厚荚相思各家系寒害率和寒害指数与树高、地径和冠幅的生长没有明显的相关关系,这与何贵平等[11]在杉木苗期抗寒性研究的结果相类似,在花旗松不同家系种苗性状变异的研究中也发现有类似的现象[16]。可能是物种之间的遗传差异所致,本文的调查分析结果说明了厚荚相思的生长与抗寒性表现为独立遗传关系。为此,在厚荚相思优良家系选择时,可以独立制定生长和抗寒性的选择指标。
本文通过田间寒害调查可为评价厚荚相思各家系的抗寒能力强弱提供参考依据,要更深入了解厚荚相思各家系间本身的抗寒特性,仍需进一步从植株在低温胁迫过程中的生理生化特性的变化深入研究探讨,从其内部生理生化机制上研究各家系的抗寒本质,从而制定出能正确反映厚荚相思各家系抗寒能力强弱的评价指标系统。
[1]潘志刚,游应天.厚荚相思的引种及种源试验[J].林业科学研究,1994,7(5):498-504.
[2]龚木荣,李忠正.值得大力推广的造纸速生材-相思木[J].中华纸业,2001,22(12):42-44.
[3]陆道调,吴保国,王希群,等.相思树种研究发展综述[J].福建林学院学报,2004,24(1):92-96.
[4]杨民权.主要热带相思在华南地区的生长及适应性探讨[J].林业科学研究,1990,3(2):155-161.
[5]韦增建,丘小军,莫钊志.相思类树种种质资源收集保存研究[J].广西林业科学,1996,25(4):81-188.
[6]续九如.林木数量遗传学[M].北京:高等教育出版社,2006:16-41.
[7]顾万春.林业试验统计[M].南宁:广西人民出版社,1984.
[8]韩宏伟,张世红,徐兴兴,等.中国刺槐种源间抗寒性地理变异研究[J].河北农业大学学报,2008,3(2):57-60.
[9]王家源,郭杰,喻方圆.不同产地苦楝苗木抗寒性差异[J].林业科技开发,2011,25(3):67-69.
[10]张卫华,张方秋,陈祖旭,等.厚荚相思抗寒性选择研究[J].广东林业科技,2009,25(5):8-13.
[11]何贵平,陈益泰,李恭学.杉木苗期抗寒性的遗传变异和家系选择[J].林业科学研究,1990,3(6):642-648.
[12]史清华,高建社,王军,等.5个杨树无性系抗寒性的测定与评价[J].西北植物学报,2003,23(11):1937-1941.
[13]卞禄,谢宝东,吴峰.不同香樟单株含水量及抗寒性差异分析[J].林业科技开发,2009,23(3):77-79.
[14]沈洪波,陈学森,张艳敏.果树抗寒性的遗传与育种研究进展[J].果树学报,2002,19(5):292-297.
[15]邹小兴,梁一池,胡彦师.巨桉种源-家系性状相关分析[J].江西农业大学学报:自然科学版,2003,25(1):60-64.
[16]张存旭,叶宏谋,韩哲.花旗松不同家系种苗性状变异分析[J].河北林果研究,1999,14(3):204-207.
猜你喜欢
今日农业(2021年20期)2021-11-26
河北果树(2020年4期)2020-11-26
大众科学(2020年6期)2020-09-06
中华建设(2019年6期)2019-07-19
沉积与特提斯地质(2019年1期)2019-07-16
乡村地理(2017年4期)2017-09-18
新疆农垦科技(2016年2期)2016-08-21
广西林业科学(2016年2期)2016-03-20
中国果菜(2016年9期)2016-03-01
中国马铃薯(2015年5期)2016-01-09
