
松嫩草原优势植物羊草和芦苇凋落物的分解对模拟增温及氮沉降的响应
2012-09-15 04:29郭继勋
东北师大学报(自然科学版) 2012年4期
齐 红,刘 洋,郭继勋
(1.长春教育学院生物系,吉林 长春 130061;2.东北师范大学草地科学研究所,吉林 长春 130024)
近年来,由于化石燃料的燃烧、不合理的农田施肥以及土地利用方式的改变,导致了温室气体(CO2,NOx,CH4)排放增加,引起的全球变暖和大气氮沉降问题已成为科学界最为关注的环境问题[1-2].温度升高几乎可影响植物所有的生物学过程,氮沉降会直接影响生物圈的氮循环,并且通过氮输入的增加影响植物生产力和生态系统碳蓄积能力,从而使生态系统结构和功能发生改变[3].
草原作为陆地生态系统的主体类型,占陆地生态系统总面积的16.4%,草地生态系统的研究是认识增温、氮沉降条件下全球生态系统变化的关键之一[4].而凋落物分解作用控制着草原生态系统养分的周转速率,与光合作用一样是养分循环的速度调节器.因此,凋落物的分解作用又是草原生态系统结构和功能研究中最重要的一个方面.
早在1876年,德国的Ebermayaer就开始研究凋落物在养分循环中的作用,此后国内外许多学者对世界范围内凋落物的分解及养分释放进行了大量报道.目前,凋落物的分解研究主要集中在两方面:一方面在继续探讨分解时的养分动态、分解与凋落物质量、生境、生物以及气候之间的关系[5-6];另一方面又将分解与一些环境热点问题结合起来加以研究,如多样性丧失对凋落物分解过程的影响,以及凋落物分解在碳循环和氮循环中的作用等[7-10].近年来,国内外许多学者对模拟增温及氮沉降条件下的凋落物分解作用进行了研究[11-12],取得了一些进展,但研究多集中在森林生态系统中,而对草地生态系统的研究较少;对于全球气候变化情况下,凋落物分解机制及微生物作用的变化还不清楚.本文在松嫩草原上进行了模拟增温和氮沉降控制实验,通过测定优势种羊草、芦苇凋落物的分解速率,凋落物中细菌、真菌、放线菌的生物量,以及凋落物养分含量的动态变化,揭示了增温及氮沉降条件下凋落物分解的变化规律,对进一步研究凋落物分解对全球气候变化下的响应机制提供了理论参考.
1 材料与方法
1.1 样地概况
实验样地位于东北松嫩草原西南部、吉林省长岭县境内(44°45′N,123°45′E).该地区为典型的大陆性气候,年均气温4.9℃;年均降雨量470.6mm,多集中在6—9月;春季多风干旱,年蒸发量为1600.2mm,约为降雨量的3.5倍.区内草原属草甸草原类型,地带性植被为草甸草原,群落类型以羊草(Leymus chinensis)+杂类草群落为主.研究样地内羊草占绝对优势,同时伴有芦苇(Phragmites communis)及多种杂类草.样地所处地段的土壤pH值为8.1左右,有机质含量为3%~4%.
1.2 实验设计
2006,2007年两个生长季,在松嫩草地选取地势平坦、植被均匀、具有代表性的地段,设置样地144m2,划分成12个(4m×3m)小区,小区间隔3m.温度控制采用红外线加热仪(infrared radiator,USA),灯管方向为南北方向,灯管距地面2.25m,温度处理保持终年进行,增温幅度为(1.7±0.1)℃.每个小区一半施氮(10g/m2),一半不施氮,氮肥采用分析纯硝酸铵.共设4个处理,由增温和施氮两个因素各两个水平组成,即增温(W)、施氮(N)、增温+施氮(WN)、对照(CK),每个处理6次重复.
1.3 实验方法
(1)凋落物的分解.凋落物分解测定采用网袋法:将优势物种羊草和芦苇的枯死体剪成约20cm的小段,并装入网眼为2mm×2mm的尼龙网袋内,每袋10g,然后将网袋放入增温、施氮处理样地.在2006和2007两年的生长季内,每隔30d取样.
(2)凋落物有机碳测定.采用重铬酸钾法:在油浴加热的条件下,用重铬酸钾-硫酸溶液来氧化凋落物中的有机碳,过量的重铬酸钾用硫酸亚铁滴定,以邻啡罗啉为指示剂,通过所消耗的重铬酸钾量计算有机碳的含量.
(3)凋落物含氮量测定.采用Foss公司生产的凯氏定氮仪测定.
(4)凋落物含磷量测定.采用端视电感耦合等离子体发射光谱法(ICP-AES)测定凋落物中的全磷含量.
(5)微生物生物量测定.凋落物微生物生物量的测定采用干重换算法:细菌生物量计算公式:

式中:B为生物量,c为细菌纯培养悬液的总干重(g),b为细菌纯培养悬液中的总数(个),N为1g凋落物中的细菌总数(个).
放线菌和真菌生物量计算公式:

式中:B为生物量,dw为5mL悬液菌丝的总干重(g),L1为5mL悬液菌丝涂片显微镜视野中菌丝的平均长度(μm),L2为50mL凋落物稀释悬液涂片中菌丝的平均长度(μm),ds为干土百分比,10为50mL稀释凋落物悬液与5mL菌丝悬液的比值.
测定的各指标均为两个生长季的平均值.
1.4 数据分析
采用SPSS14.0软件进行统计分析,采用Sigmaplot10.0软件绘图.
2 结果
2.1 凋落物分解对增温、施氮的响应
羊草和芦苇是松嫩草原典型的优势种,但两者的营养组成成分差别很大,羊草中富含有机质,而芦苇中含大量的纤维木质素,因此,羊草凋落物的分解速率要快于芦苇凋落物.
羊草凋落物经各处理分解后,残留率从小到大依次为:增温+施氮处理69.30%,施氮处理75.75%,增温处理81.20%,对照83.80%.增温和施氮处理均能增加羊草凋落物的分解速率,二者交互作用不显著(见图1).
芦苇凋落物经各处理分解后,残留率从小到大依次为:增温+施氮处理78.85%,施氮处理83.25%,对照85.60%,增温处理86.50%.其中增温处理对芦苇凋落物分解无显著作用,施氮处理显著提高了芦苇凋落物的分解作用.而增温+施氮处理对芦苇凋落物分解速率有促进作用,超过增温和施氮处理之和,呈显著的正交互作用(见图2).
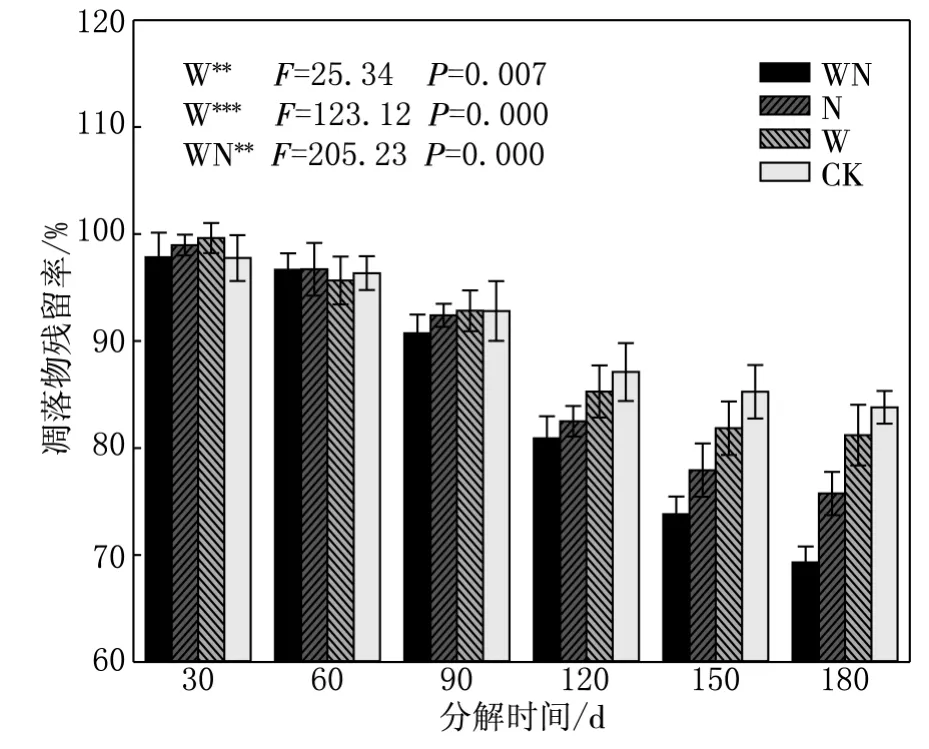
图1 羊草凋落物量季节动态

图2 芦苇凋落物量季节动态
采用Olson(1963)负指数方程来模拟分解过程[13]:

式中:t为分解时间(d);Wt为时间t时凋落物残余的质量,W0是起始时凋落物的质量;k是凋落物质量丢失的速率系数.分解周期指95%的凋落物被分解所需要的时间,利用k值计算在4种处理下羊草和芦苇凋落物分解周期.
负指数模型对羊草和芦苇凋落物分解的拟合度均达到R2>0.9.在平均地表增温1.7℃,每个生长季施氮(10g/m2)的情况下,羊草和芦苇分解周期都有不同程度的减少,特别是增温+施氮处理下的羊草凋落物分解周期缩短了49.98%(见表1).这将极大加快羊草草原的碳、氮循环周期.
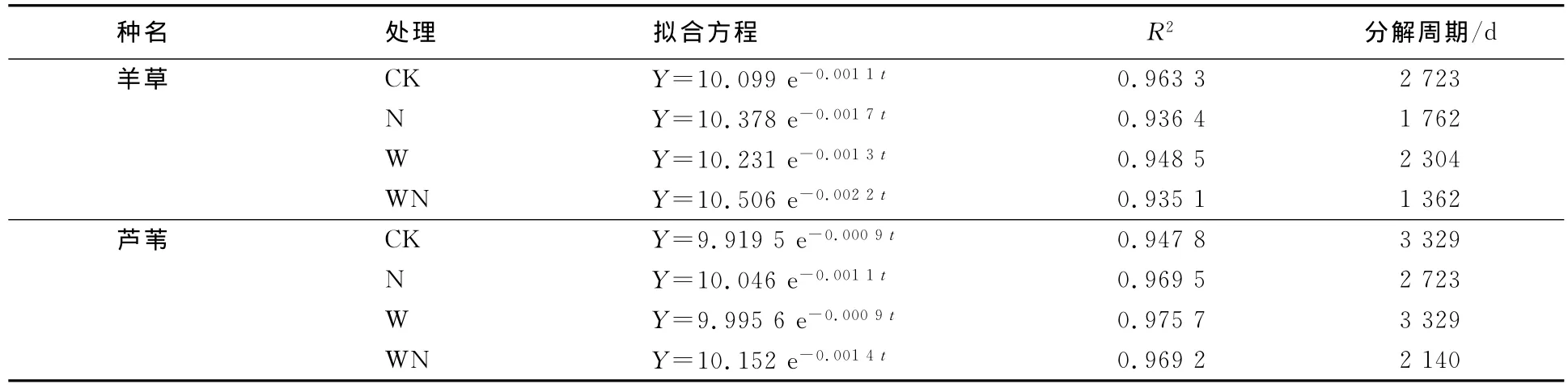
表1 凋落物剩余量与时间关系的拟合
2.2 微生物生物量对增温、施氮的响应
在草原生态系统中,凋落物的分解主要靠微生物来完成,其分解强度占枯枝落叶年损失率的97%[32].羊草凋落物在增温处理下细菌、真菌和放线菌生物量分别增加了43.51%,4.69%,58.28%;而在施氮处理下细菌、真菌生物量分别减少了9.74%和30.82%,放线菌增加了19.64%(见图3).
芦苇凋落物在增温处理后细菌生物量减少了15.15%,真菌和放线菌生物量分别增加了38.80%,289.47%;施氮处理后细菌、真菌和放线菌生物量分别增加了24.24%,28.91%,185.71%(见图4).
以4种处理下分解的羊草与芦苇凋落生物量为因变量,凋落物中细菌、真菌、放线菌的生物量为自变量做多元回归分析,结果见表2.回归分析表明,细菌、真菌、放线菌生物量均不同程度地受到增温和施氮的影响.两种凋落物各种处理之间的变化结果基本一致,所有方程中细菌的标准化系数均为最大,即表示在增温和施氮处理下,羊草与芦苇的细菌生物量始终对凋落物分解的贡献最大,真菌和放线菌相对较弱.增温处理减少了细菌的贡献,却显著增加了真菌对分解速率的影响;施氮处理在降低了细菌作用的同时却增加了真菌和放线菌对凋落物分解的贡献.增温+施氮处理显著的增加了3种菌类对凋落物分解的贡献.

图3 羊草凋落物三种微生物生物量
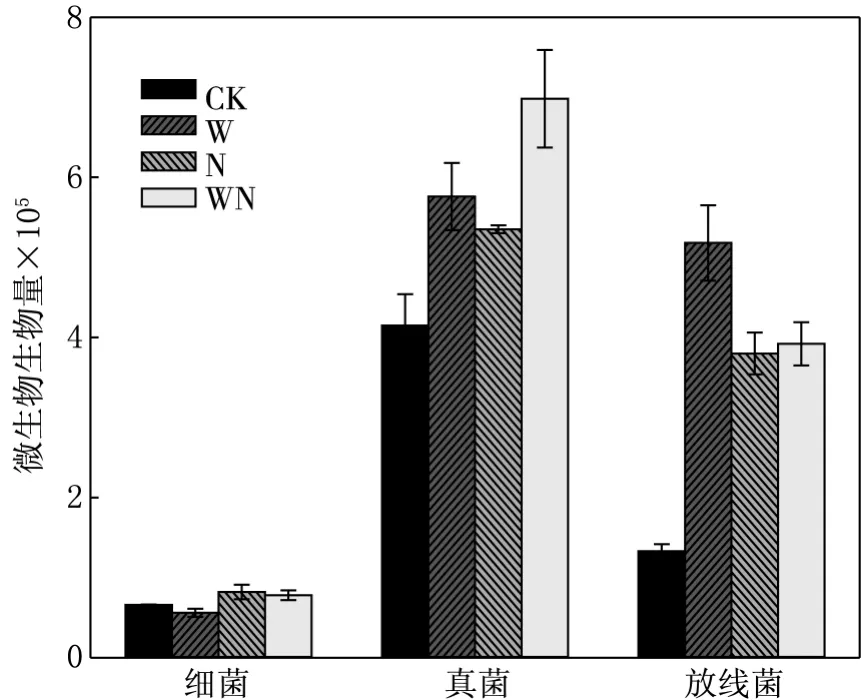
图4 芦苇凋落物三种微生物生物量

表2 真菌、细菌、放线菌与凋落物分解量的多元回归分析
2.3 凋落物分解过程中C,N,P的变化
羊草凋落物分解过程中,增温使凋落物的N,P含量分别增加了20.86%,19.27%,使有机C含量、C/N降低了59.85%,66.88%;施氮使N,P含量分别增加28.52%,31.81%,使有机C含量、C/N降低了为58.45%,67.68%;增温+施氮处理使N,P含量增加了16.92%,21.17%,使有机C含量、C/N降低了90.58%,91.95%.
芦苇凋落物分解过程中,增温使N,P含量分别增加12.64%,32.12%,使有机C含量、C/N降低了60.51%,64.95%;施氮使N,P含量分别增加20.25%,30.56%,使有机C含量、C/N降低了76.51%,80.47%;增温+施氮处理使N,P含量增加30.67%,37.21%,使有机C含量、C/N降低了71.16%,75.58%(见表3).两种植物凋落物分解过程中都表现为有机C含量迅速减少、凋落物中的N,P含量固持的现象.

表3 羊草和芦苇凋落物中的C,N,P分析
3 讨论
在全球气候变化的综合作用下,凋落物分解速率的加快是不容置疑的.已有研究表明,氮沉降使氮的输入增加,极大地促进了凋落物CO2和NO2的呼吸过程,促进了凋落物的分解,而且在凋落物分解最初阶段,分解的对象是一些易分解的非木质性的纤维素和有机碳,这些易分解的物质是微生物所利用的主要碳源.凋落物属于普遍氮缺乏型,而外源氮的输入恰恰满足了微生物对氮的需求,刺激了微生物的活动,因而加速了分解过程[14-15].这些观点解释了我们在施氮处理中羊草和芦苇凋落物分解速率加快、分解周期缩短及有机碳大量释放的结果.但也有研究表明,残留物中的木质素和全纤维素质量分数随着外源氮梯度增加均显著增加,而且在长期高浓度氮存在的情况下,氮可能与凋落物分解过程中的某些物质聚合,形成更难降解的物质,从而阻滞了凋落物的分解[19].
大多研究结果认为:增温会加快分解速率.Silver和Miya总结了全球细根植物分解数据时发现,年平均温度与凋落物分解率呈线性正相关[20];Vitousek等在太平洋热带岛屿Mauna Loa的研究表明,随海拔升高及气温降低,凋落物的分解速率呈指数降低[6].这些结果支持了我们增温处理中羊草凋落物分解速率增加、分解周期缩短的结论.
然而,在我们的结果中,增温并没有显著提高芦苇凋落物的分解速率,这可能与C/N的变化有关.增温处理显著地提高了芦苇凋落物C/N的比值,由于C/N的比值较大,氮素的缺乏使微生物的发育受到了限制,从而使有机质分解变得缓慢.同时,凋落物的细菌、真菌、放线菌群落结构的变化也会对分解速率产生影响,与对照相比,增温处理下芦苇凋落物分解量与微生物生物量的回归分析表明:3种微生物对分解量的贡献都有所降低(见表3).
对于羊草来说,增温使3种菌类的生物量都有所提高,而施氮使细菌和真菌的生物量减少,这说明,氮元素不是制约分解速率的关键因素.而对于芦苇来说,恰恰相反,施氮样地凋落物中3种菌类生物量都有所增加,而增温样地增加却不明显,氮元素是微生物生长的限制因素.
羊草和芦苇凋落物中的N,P元素含量在生长季结束的时候都比生长季前有所提高,这与许多研究结果相同.通常认为凋落物N,P元素含量升高的固持现象主要发生在第一年分解时间内,之后即进入矿化阶段[21].在本实验中,所有样地都出现凋落物N,P元素含量升高的固持现象,生长季结束后N,P元素含量和对照比较均达到了极为显著的水平;对于增温+施氮处理的样地,这种固持现象更加明显.固持现象可能是由于凋落物中微生物大量生长,而微生物的这种生长需要大量的N和P来构成身体的组成部分,所以微生物不仅从凋落物中吸收N,P,而且要从凋落物以外的环境中吸收N,P,从而使N,P元素在凋落物中聚集,使凋落物在分解初期N,P元素含量升高.一直到凋落物中的其他元素(例如有机碳、K)被微生物呼吸、或被土壤淋溶耗尽后,凋落物中的N,P再进行矿化作用,释放到环境中去.从宏观的角度说:凋落物层具有改善草原生态系统的作用可能也与这种N,P元素的固持现象有关,而增温及氮沉降对这种固持现象有较明显的促进作用.[22]
[1]BURTON D L,BEAUCHAMP E G.Profile mitrours oxide and carbon dioxide concentrations in a soil subject to freezing[J].Soil Sci Soc,1994,58:15-122.
[2]CHRISTENSEN T R,JONASSON S,CALLAGHAN T V,et al.On the potential CO2release from tundra soils in a changing climate[J].Applied Soil Ecology,1999,11:127-134.
[3]吕超群,田汉勤,黄 耀.陆地生态系统氮沉降增加的生态效应[J].植物生态学报,2007,31(2):205-218.
[4]OJIMA D S,PARTON W J,SCHIMEL D S,et al.Modeling the effects of climate and CO2changes on grassland storage of soil C [J].Water Air Soil Pollution,1993,70:643-657.
[5]HOBBIE S E.Temperature and plant species control over litter decomposition in Alaskan Tundra[J].Ecological Monographs,1996,66(4):503-522.
[6]VITOUSEK P M,TURNER D R,PARTON W J,et al.Litter decomposition on the Mauna Loa environmental matrix,Hawaii:patterns,mechanisms,and models[J].Ecology,1994,75(2):418-429.
[7]AERTS R.The freezer defrosting:global warming and litter decomposition rates in cold biomes[J].Journal of Ecology,2006,94(4):713-724.
[8]莫江明,薛碌花,方运霆.鼎湖山主要森林植物凋落物分解及其对 N沉降的响应[J].生态学报,2004,24(7):1413-1419.
[9]DUKES J S,FIELD C B.Diverse mechanisms for CO2effects on grassland litter decomposition[J].Global Change Biology,2000,2(6):145-154.
[10]XIAO D M,WANG M,JI L Z,et al.Variation characteristics of soil N2O emission flux in broad-leaved Korean pine forest of Changbai Mountain[J].Chinese Journal of Ecology,2004,23(5):46-52.
[11]陈华,MARK E HARMONL,田汉勤.全球变化对陆地生态系统枯落物分解的影响[J].生态学报,2001,21(9):1549-1563.
[12]HOBBIE S,GOUGH L.Litter decomposition in moist acidic and non-acidic tundra with different glacial histories[J].Oecologia,2004,140:113-124.
[13]MELILLO J M,ABER J D,MURATORE J F.Nitrogen and lignin control of hardwood leaf litter decomposition dynamics[J].Ecology,1982,63:621-626.
[14]ANDERSON J M,HETHERINGTON S L.Temperature,nitrogen availability and mixture effects on the decomposition of heather and bracken litters[J].Functional Ecology,1999,13:116-124.
[18]MICKS P,DOWNS M R,MAGILL A H.Decomposition litter as a sink for 15N2enriched additions to an oak forest and a red pine plantation[J].Forest Ecosystem and Management,2004,196:71-87.
[19]LIMPENS L,BERENDSE L.How litter quality affects mass loss and N loss from decomposing Sphagnum[J].Oikos,2003,103:537-547.
[20]SILVER W L,MIYA R K.Global patterns in root decomposition:comparisons of climate and litter quality effects[J].Oecologia,2001,129:407-419.
[21]JOSEPH J H,WILSON C A,BORINGL R.Foliar litter position and decomposition in a fire-maintained longleaf pine-wiregrass ecosystem[J].Canadian Journal of Forest Research,2002,32:928-941.
[22]刘洋.松嫩草地优势植物羊草、芦苇凋落物分解对模拟增温及施氮的响应[D].长春:东北师范大学,2008.
猜你喜欢
农业科技通讯(2023年1期)2023-02-12
现代畜牧科技(2021年7期)2021-07-28
现代畜牧科技(2021年4期)2021-07-21
国际呼吸杂志(2019年22期)2019-12-09
农业环境科学学报(2017年2期)2017-03-20
现代畜牧科技(2016年9期)2016-02-21
草业科学(2015年9期)2015-12-11
应用海洋学学报(2014年4期)2014-11-22
农家顾问(2014年9期)2014-10-20
天然产物研究与开发(2014年6期)2014-04-27
