
红菜薹细胞质雄性不育系的农艺性状调查及POD同工酶分析
2012-08-07 09:43:30邵明珠徐跃进万正杰邓霜
中国蔬菜 2012年2期
邵明珠徐跃进万正杰邓霜
(华中农业大学园艺林学学院,华中农业大学园艺植物生物学教育部重点实验室,湖北武汉430070)
国内外对十字花科中的甘蓝型油菜雄性不育的研究和利用较为广泛(Shiga & Baba,1973;刘忠松和官春云,1990;Wang et al.,2000)。王亚馥等(1984)比较研究了大白菜雄性不育两用系可育株和不育株花苞的过氧化物同工酶,结果表明不育株的过氧化物酶活性高于可育株,而且谱带多。梁燕等(1994)以大白菜为试材,进行过氧化物同工酶研究,也得到了与王亚馥(1984)相似的结论。许忠明等(2004)以甘蓝胞质雄性不育系及其保持系为试材,分析了POD同工酶的谱带差异。结果表明,不育系和保持系间POD同工酶差异以花、大蕾和小蕾最大,叶和根间差异不明显。在花和花蕾中,不育系酶带活性强于相应的保持系,在根中正好相反。但关于红菜薹〔Brassica campestrisL. ssp.chinensis(L.)Makino var.utilisTsen et Lee〕胞质雄性不育系和保持系不同组织器官POD同工酶的变化规律以及POD同工酶与红菜薹胞质雄性不育的关系目前鲜有报道。
本试验利用新型甘蓝型油菜雄性不育材料(EruCMS)细胞质雄性不育系与红菜薹保持系俞优杂交后,再经过连续的多代回交,获得了红菜薹异源细胞质雄性不育系材料,其不育率100%,低温不黄化,蜜腺正常,同时以红菜薹保持系俞优为材料研究不同器官的POD同工酶的变化,探讨红菜薹细胞质雄性不育形成的机理。
1 材料与方法
1.1 材料
新型甘蓝型油菜雄性不育材料(EruCMS)细胞质雄性不育系(用油菜P系表示)与红菜薹保持系俞优(用红菜薹B系表示)杂交后,再经过连续的多代回交,获得了红菜薹异源细胞质雄性不育系材料(用红菜薹A系表示)。
1.2 方法
1.2.1 油菜P系、红菜薹B系和红菜薹A系的形态观测 试验于2010年在华中农业大学蔬菜实习基地大棚进行,试材于9月25日定植于大棚,常规田间管理。10月24日用卷尺和游标卡尺测定油菜P系、红菜薹B系和红菜薹A系3种试材叶片生长期植株的株高、开展度、叶柄的长和粗、最大叶片的长和宽等形态特征;11月25日测定3种试材的花朵开展度、花瓣的长和宽、花柱长、雄蕊长、花丝长等性状,每种试材测10株,3次重复,然后对3种试材花器官拍照并采用TTC法(王学奎,2006)测定其花粉活力。
1.2.2 油菜P系、红菜薹B系和红菜薹A系的POD同工酶分析 在不育系和保持系群体中随机取样,分别取其叶片、叶柄、薹、花蕾(大蕾≥3.5 mm,中蕾2~3 mm,小蕾<1 mm)和当日开放的花为试样。电泳采用聚丙烯酰胺凝胶垂直板电泳,配方参考郭尧君(1999)的方法,同工酶染色方法采用联苯胺染色法(许忠民 等,2004)。

2 结果与分析
2.1 油菜P系、红菜薹B系和红菜薹A系外部形态和特性的比较
3种试材的花器官外部形态和特性见图1,油菜P系、红菜薹B系和红菜薹A系花器官外部特征基本相同,自外向内分别由4枚萼片、4枚花瓣排成十字型,雄蕊6枚,四强雄蕊,外轮2枚较短,内轮4枚较长(图1-P1、A1、B1);红菜薹B系的花药基本和柱头等高,雄蕊有花粉,有利于正常的授粉结实(图1-B2、B3)。油菜P系花药直立,紧贴柱头,但远远低于柱头(图1-P2、P3),而红菜薹A系的花药向四周伸展,但二者蜜腺都正常,雌蕊都能正常的授粉结实,而且不受温度的影响,不育度和不育率达100%(图1-A2、A3)。
2.2 油菜P系、红菜薹B系和红菜薹A系农艺性状的调查分析
通过对油菜P系、红菜薹A系和红菜薹B系在生长期的形态指标和花器官性状作比较得出(表1):红菜薹B系生长期的最大叶片宽显著大于油菜P系和红菜薹A系,而最大叶片长三者之间差异不显著;从三者的株高上看,油菜P系的最高,红菜薹A系次之,而红菜薹B系最矮;红菜薹A系与油菜P系的叶柄长和粗达到了显著差异水平;植株开展度三者之间没有显著差异。

图1 油菜P系、红菜薹B系与红菜薹A系的花器官外部形态和特性
油菜 P系、红菜薹 A系和红菜薹 B系在花器官性状方面存在差异,有的甚至达到了显著或极显著水平。在花朵的开展度方面,三者间红菜薹 A系的最小,而油菜 P系、红菜薹B系之间没有显著差异;在花瓣大小方面,红菜薹A系的花瓣长和宽最小,与油菜 P系、红菜薹 B系之间存在极显著差异;油菜 P系的花柱最长,比红菜薹 A系和红菜薹B系材料高出19%、18%,而红菜薹 A系和红菜薹 B系的花柱长没有显著差异;但在雄蕊长、花丝长以及雌雄蕊高度差等方面油菜P系、红菜薹A系和红菜薹B系均达到了极显著差异,且红菜薹 A系居于二者之间,中和了父母本性状,充分体现了杂种优势。
2.3 油菜P系、红菜薹B系和红菜薹A系的花粉活力分析
从2010年11月~2011年3月上旬连续对油菜P系、红菜薹A系和红菜薹B系的花粉进行活力检测。红菜薹B系的花粉被TTC染成了红色(图2-B),说明花粉具有活力;油菜P系(图2-P)和红菜薹A系(图2-A)的花粉被染成浅黄色,说明花粉无活力,红菜薹A系育性很稳定。
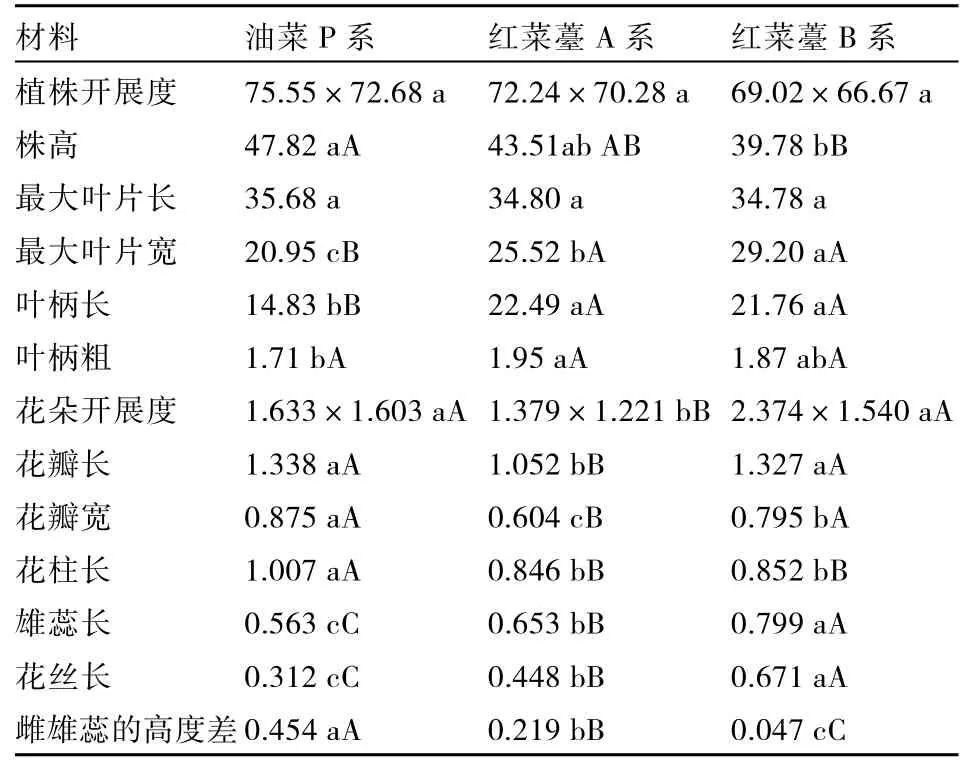
表1 油菜P系、红菜薹B系和红菜薹A系农艺性状的调查分析结果 cm
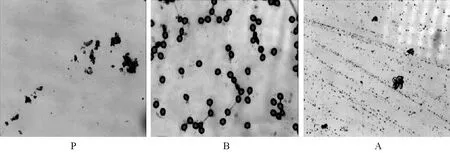
图2 油菜P系、红菜薹B系和红菜薹A系的花粉活力(×10倍)
2.4 油菜P系、红菜薹B系和红菜薹A系的POD同工酶分析
油菜P系、红菜薹A系和红菜薹B系材料从叶片、叶柄、薹3个部位的POD同工酶酶谱来看(图3、4):负极到正极出现了1~5条谱带,其中Rf=0.409是一条共有谱带;红菜薹B系和油菜P系叶片的谱带比红菜薹A系多,油菜P系特有Rf=0.606和0.682的谱带,而红菜薹B系特有Rf=0.227的谱带;红菜薹A系和红菜薹B系叶柄的谱带基本没区别,与油菜P系仅有一条共有谱带;在薹中从负极到正极,POD同工酶的谱带有3条或5条,红菜薹A系比红菜薹B系少两条弱谱带(Rf=0.272和0.788)。
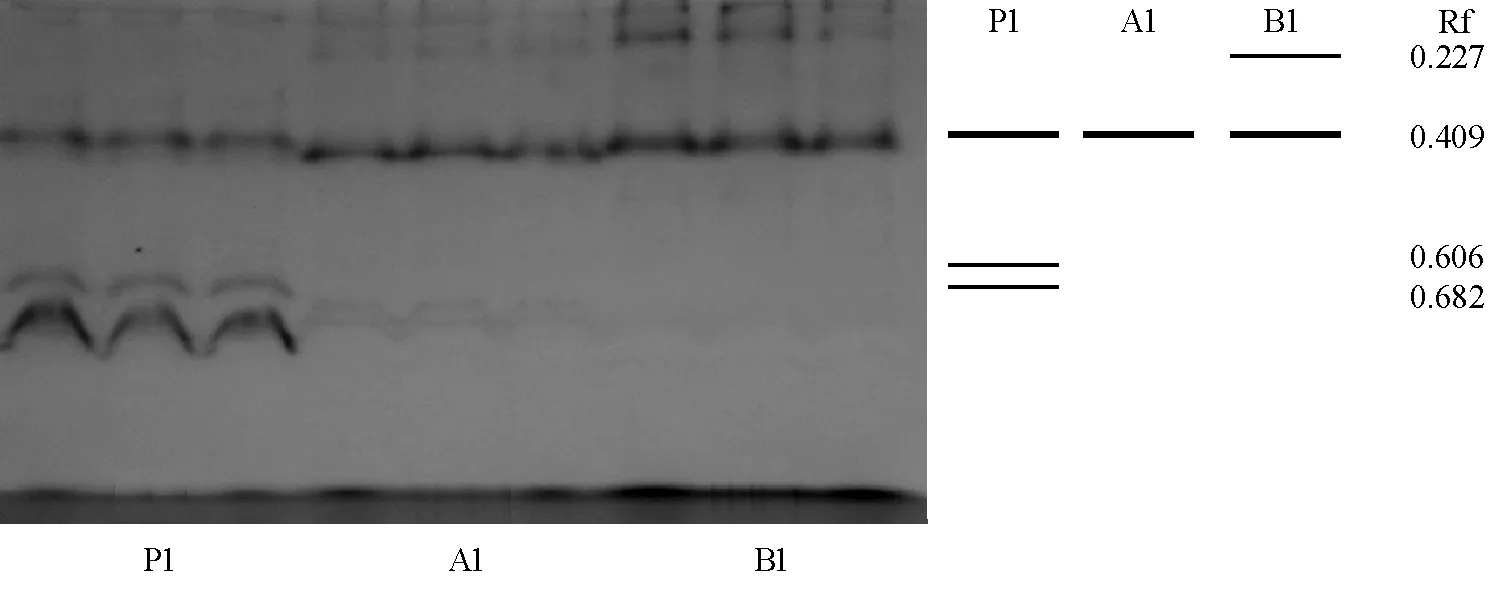
图3 油菜P系、红菜薹B系和红菜薹A系叶片POD同工酶酶谱和模式图

图4 油菜P系、红菜薹B系和红菜薹A系叶柄和薹POD同工酶酶谱和模式图
从油菜P系、红菜薹A系和红菜薹B系材料花和不同级别花蕾4种试样POD同工酶酶谱可以看出(图5):由负极到正极出现了1~4条谱带,Rf=0.446是一条共有谱带,但不同类型试材和同一试材不同器官间酶谱带存在强弱差异性,说明此条带在表达上具有很强的阶段性;三者花(P4、A4、B4)中具有相同的POD同工酶谱带,而不同级别的蕾中POD同工酶谱带存在差异;在油菜P系的小蕾(P7)中比其他试材(A7、B7)多两条弱谱带,红菜薹A系的中蕾(A6)比其他两份材料(P6、B6)多一条Rf=0.257的谱带,是其特有的一条弱的谱带,有可能与其不育相关。

图5 油菜P系、红菜薹B系和红菜薹A系盛花期POD同工酶酶谱差异表现
由3类试材的POD同工酶酶谱比较可以看出,油菜P系、红菜薹A系在叶片、叶柄及蕾的POD同工酶酶谱差异较大,而红菜薹A系和红菜薹B系在叶柄中的POD同工酶酶谱没有区别,而在叶片、薹及蕾中有明显差别,特别是红菜薹A系和红菜薹B系蕾由小到大,同工酶POD谱带上则表现出由少到多,谱带强弱差异较大。从花药到小孢子的发育阶段来看,可能是花粉败育之前,物质合成代谢旺盛,POD活性相对较弱。花粉开始败育后,物质分解代谢加强,POD同工酶谱带上则表现出谱带多,染色深。
3 结论与讨论
3.1 红菜薹异源胞质雄性不育系(红菜薹A系)、红菜薹自交系(红菜薹B系)与新型甘蓝型油菜雄性不育材料(Eru CMS)(油菜P系)不育系农艺性状调查分析
植物性状表现主要由遗传基因决定,试验的观察结果表明,油菜P系、红菜薹A系和红菜薹B系3类供试材料在花器官的形态上7个主要性状之间都存在显著差异,其中在雄蕊长、花丝长、雌雄蕊的高度差方面,三者差异规律性变化是红菜薹A系居中,在花柱性状方面红菜薹A系和红菜薹B系没有区别,油菜P系最大;而在花瓣的长和宽方面,油菜P系最大、红菜薹B系居中、红菜薹A系最小。生长期的植物学性状调查显示,差异性不显著,没有规律性可寻。分析原因可能是由于这些性状多为数量基因决定的性状遗传,红菜薹A系材料是红菜薹B系与油菜 P系的杂交后代,通过多代回交核基因未纯合之前仍处于中间型,因而表现出花器大小居中的特性。由此结果也说明红菜薹自交系转育新型甘蓝型油菜雄性不育材料(EruCMS)基因通过杂交和回交是可行的,红菜薹的核基因在其回交后代中是能够表现的。
从2010年11月~2011年3月上旬连续对BC6的花粉进行活力检测,这期间武汉经历了多次寒流的袭击,气温变化剧烈,但均未观察到可育花粉粒,说明 BC6的不育性很稳定。说明转育是可行的,可以对其配制杂交组合,进行配合力的测定。
3.2 红菜薹A系、红菜薹B系与油菜P系的POD同工酶分析
对于POD同工酶与雄性不育的关系,一种观点认为雄性不育与花药(或整个生殖器官)中的膜脂过氧化作用密切相关,膜脂过氧化受体内的活性氧含量的直接影响,而过氧化物酶对清除活性氧有重要作用。但一般认为过氧化物酶同工酶基因是在雄性不育发生过程中被动开启的,即过氧化物酶同工酶活性高的时候就是活性氧含量高的时候,也就是说在幼蕾期正在发生着强烈的膜脂过氧化作用,进而引起了一系列败育过程,导致雄性不育的发生(任雪松 等,2004)。
本试验油菜P系、红菜薹A系和红菜薹B系3类供试材料的在各个器官当中的POD同工酶分析显示,红菜薹A系和红菜薹B系间的差异主要发生在生殖生长时期,在抽薹期,红菜薹A系的薹比红菜薹B系的薹少了两条谱带(Rf=0.272和Rf=0.788);在特定的蕾期差异比较明显,而且红菜薹A系比红菜薹B系在中蕾中多了1条谱带(Rf=0.257),这与梁燕等(1994)、张丽等(2006)及赵会芳等(2009)的研究结果一致,与刘忠松和官春云(1990)的研究结果相反。刘忠松和官春云(1990)研究指出:开花期花蕾中,不育系较保持系的过氧化物酶谱带少、酶活性弱,随着花蕾的发育,保持系增加了1条谱带,且酶活性亦有增加趋势。由于甘蓝型油菜、萝卜、大白菜和甘蓝均为十字花科的作物,有一定的可比性。可见,这些细胞质雄性不育的过氧化物酶同工酶在生殖器官(花蕾)中存在差异这一点上是一致的,但具体差异的形式和程度却因植物而异。
郭尧君.1999.蛋白质电泳技术.北京:科学出版社:70-73.
梁燕,王鸣,赵稚雅.1994.结球白菜同核异质雄性不育系同工酶研究.园艺学报,21(3):264-268.
刘忠松,官春云.1990.油菜雄性不育性的研究——Ⅰ.甘蓝型油菜波里马(Polima)细胞质雄性不育系与保持系的生化比较.中国油料,12(3):1-4.
任雪松,李成琼,宋洪元.2004.甘蓝胞质雄性不育系和保持系花药同工酶分析.西南农业大学学报:自然科学版,26(4):433-436.
王学奎.2006.植物生理生化实验原理和技术.2版.北京:高等教育出版社:220-221.
王亚馥,胡昌勤,安旺盛.1984.大白菜(Brassica pekinensis)雄性可育株与雄性不育株花苞的核酸,蛋白质,酶的初步分析.兰州大学学报:自然科学版,20(1):90-96.
许忠民,巩振辉,王妍妮,张恩慧,程永安.2004.甘蓝胞质雄性不育系和保持系POD同工酶分析.西北植物学报,24(1):21-24.
赵会芳,巩振辉,赵利民,柯桂兰.2009.大白菜萝卜细胞质雄性不育系 RC7花药发育的解剖学和同工酶研究.中国农业大学学报,14(2):64-69.
张丽,宫国义,李霄燕,陈静华.2006.萝卜雄性不育系及其保持系个体发育中5种同工酶比较.西北农业学报,15(3):116-120.
Shiga T,Baba S.1973.Cytoplasmic male sterility in oilseed rapeBrassica napusL.and its utilization to breeding.Breeding,22:187-197.
Wang Junxia,Yang Guangsheng,Fu Tingtong,Meng Jingling.2000.Development of PCR-based markers linked to the fertility restorer gene for the polinma cytoplasmic male sterility in rapes seed(Brassica napusL.).Acta Genetica Sinica,27(11):1012-1017.
猜你喜欢
遗传(2023年8期)2023-08-25 06:59:00
园艺与种苗(2021年10期)2021-11-28 23:31:51
宁夏医学杂志(2020年3期)2021-01-21 08:23:38
农药科学与管理(2020年3期)2020-07-30 01:32:46
广西林业科学(2019年2期)2019-07-25 02:29:00
中国种业(2018年8期)2018-08-28 06:40:46
东北林业大学学报(2016年12期)2017-01-09 07:41:57
种子科技(2015年8期)2015-05-24 16:14:56
安徽农业科学(2015年13期)2015-01-13 06:20:26
森林工程(2014年5期)2014-08-23 03:59:14
