
中国黄牛Y-STRs遗传多样性与起源研究
2012-08-04 06:49常振华黄洁萍张润锋雷初朝
中国牛业科学 2012年3期
常振华,黄洁萍,徐 苹,李 冉,张润锋,陈 宏,雷初朝*
(1.渭南职业技术学院,陕西 渭南 714000;2.西北农林科技大学动物科技学院,陕西 杨凌 712100;3.湖北师范学院生命科学学院,湖北 黄石 435002)
中国黄牛是指除牦牛和水牛以外的所有家牛,起源上包括普通牛和瘤牛两个牛种,二者的共同祖先是原牛。作为世界牛品种资源宝库的重要组成部分,我国地方黄牛品种具有丰富的遗传多样性[1]。牛在人类文明的发展和进步中发挥着重要的作用而且也是最具有经济价值的家畜之一。为了更好地分析家牛的起源和分布,人们提出了不同的理论。现在普遍认同的“双重驯化”观点认为,由于气候环境和牛群生理特征的不同,普通牛起源于新月形地区,而具有肩峰的瘤牛则起源于干旱的印度次大陆[2]。
中国黄牛品种资源十分丰富,适应性强,肉质良好,特别是黄河中、下游流域的许多品种或群体经过长时间的选育,其肉用性能更好。我国黄牛足迹遍布全国,分布于我国的29个省、自治区和直辖市,根据地域可以分为北方黄牛、南方黄牛和中原黄牛三大类[3]。
国内外动物遗传学家一直对中国黄牛的起源进化与遗传多样性的课题有着浓厚的兴趣。目前认为,我国黄牛为多元起源的,但究竟于何时何地起源于哪几个牛种,观点各不相同。自20世纪80年代以来,众多学者对我国地方黄牛的染色体核型进行了分析,指出不同黄牛品种的Y染色体形态具有明显的多态性,包括中部(亚中部)着丝粒和端部着丝粒Y染色体,从而陈宏等[4]指出我国地方黄牛具有普通牛和瘤牛两种血统;陈幼春等[5]认为我国地方黄牛起源于普通牛和瘤牛,同时在岭南地区和东南沿海的地方黄牛发现有印度尼西亚爪哇牛的血统。mtDNA D-loop序列分析表明,中国黄牛主要有普通牛和瘤牛两种母系起源[6,7]。动物的Y染色体遵循父系遗传,单倍型完整,不易受重组和回复突变的影响,突变率低,是进化事件的忠实记录者,是研究动物父系起源及迁徙路线的理想工具。微卫星序列(STR)是指以1~6个核苷酸为基本重复单位的串联重复序列,其长度一般在200bp以内[8]。近年来,家牛Y染色体微卫星多态性(Y-STR)研究把牛分为普通牛和瘤牛两种父系起源[9,10]。家牛Y染色体基因组测序基本完成,这样就使得许多Y染色体特异性DNA标记被鉴定,并且用于研究家牛起源、驯化及迁徙。
国内利用Y-STR标记研究黄牛Y染色体分子遗传多样性与起源进化的报道不多[11,12],因此,本研究利用2个经典的家牛Y-STR标记检测中国黄牛的Y染色体多样性,其结果为建立我国地方黄牛品种资源分子遗传多样性数据库、遗传资源保存策略与肉牛新品种培育提供科学依据。
1 材料与方法
1.1 试验动物
按照随机典型抽样的方法,本研究选择来自三大类地区的16个中国地方黄牛品种,其中北方黄牛:哈萨克牛、安西牛和蒙古牛;中原黄牛:早胜牛、晋南牛、鲁西牛、秦川牛、郏县红牛和渤海黑牛;南方黄牛:南阳牛、宣汉牛、吉安牛、海南牛、皖南牛、雷州牛和恩施牛。总计284头公牛样本,其中244头血样,40个组织样(脾脏和耳尖),另外采集缅甸黄牛4头公牛及3头秦川牛母牛的血样作对照。
1.2 方法
1.2.1 黄牛基因组DNA的提取与Y-STRs位点的选择 本研究采用常规的酚-氯仿法提取基因组DNA,并且选用了家牛2个经典的Y-STRs位点,即INRA189和BM861标记用以研究中国黄牛的父系起源,各标记的相关信息见表1,其引物由上海生工生物工程技术服务有限公司合成。

表1 家牛2个Y-STRs位点的名称、位置、重复类型、退火温度与引物序列
1.2.2 黄牛Y-STRs位点的PCR体系与非变性聚丙烯酰胺凝胶电泳多态性分析 2个黄牛Y-STRs标记的PCR扩增总体积均为12.5μL/个体,PCR扩增程序为常规程序:95℃预变性4min;94℃变性30s,59℃退火60s,72℃延伸90s,35个循环;72℃充分延伸10min,4℃保存。
PCR扩增产物通过30%非变性聚丙烯酰胺凝胶电泳分离带型,Marker为pBR322;然后预电泳10min,再用120V恒压预电泳10min后80V恒压电泳过夜;凝胶用0.1%硝酸银染色,2% 氢氧化钠和甲醛显色,用Discovery Series Quantity One Software Version4.3.1软件(Bio-Rad Laboratories Inc)进行片段大小统计和基因型分析。
2 结果与分析
2.1 中国黄牛的Y-STR多态性
本研究利用2个家牛Y染色体微卫星标记(INRA189和BM861)检测中国黄牛16个品种284头公牛和4头缅甸黄牛公牛共计288头黄牛的遗传多态性。结果表明,2个Y-STR标记INRA189和BM861在中国黄牛中各检测到3个等位基因(图1),表现出明显的多态性。在INRA189位点中,中国黄牛的3个等位基因大小分别为166、156和144 bp。其中瘤牛的等位基因大小为156/144bp,普通牛为166/156bp;在BM861位点中,中国黄牛的3个等位基因大小分别为187、169和167bp,其中瘤牛在该位点的等位基因大小为187/167bp,普通牛为187/169bp(表2和图1)。表明这2个Y-STR位点可以很好地区分中国黄牛中的普通牛与瘤牛起源。

表2 两个牛Y-STR标记的等位基因数及其变异范围
2.2 普通牛和瘤牛在中国黄牛中的分布频率
通过分析2个Y-STR位点,可以把中国黄牛分为普通牛与瘤牛两个父系起源,同时发现它们的频率在不同的品种间各不相同(表3)。总的来说,在中国16个黄牛品种中,普通牛单倍型频率为57%(162/284),高于瘤牛单倍型频率43%(126/284)。在北方牛群体中,普通牛单倍型频率占优势(98.3%),在南方牛群体中,瘤牛单倍型频率占优势(76.1%),而中原黄牛群体包括普通牛和瘤牛起源,其中普通牛单倍型频率(63.8%)高于瘤牛单倍型频率(36.2%)。
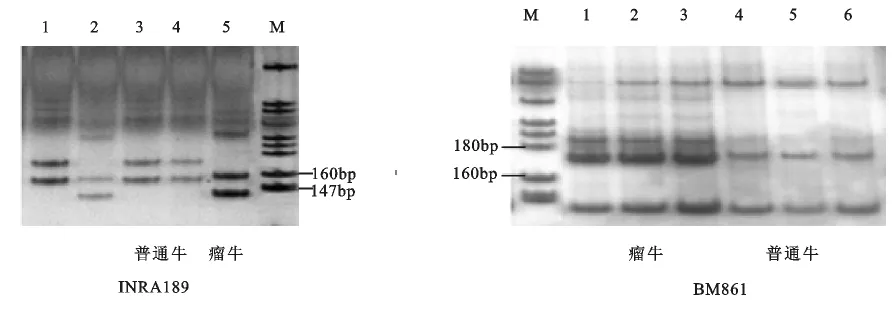
图1 黄牛2个Y-STR标记的基因分型
在北方牛群体中,蒙古牛和哈萨克牛的普通牛单倍型频率最高,均为100%,安西牛为96.4%。在南方牛群体中,宣汉牛、皖南牛、海南牛和对照组缅甸牛的瘤牛单倍型频率均为100%,而吉安牛、南阳牛、雷州牛和恩施牛的瘤牛单倍型频率分别为28.6%、29.0%、86.4%和88.9%。在中原黄牛6个品种中,其中5个品种具有普通牛和瘤牛起源。说明中国黄牛中的普通牛和瘤牛在中原地区交汇,普通牛频率自北向南逐渐减少,瘤牛频率自北向南逐渐增加。
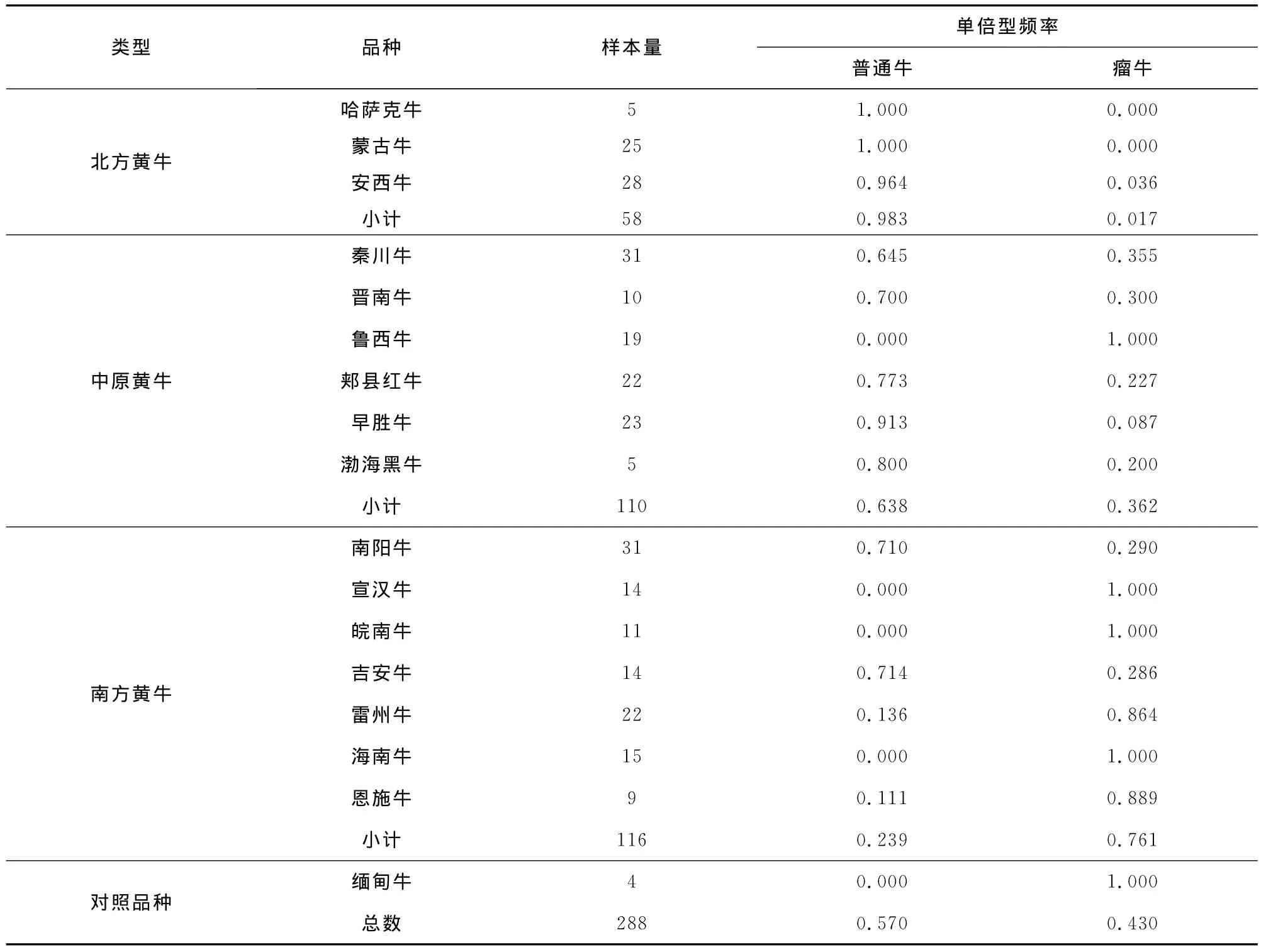
表3 中国16个地方黄牛品种和缅甸牛的普通牛和瘤牛分布频率
3 讨论
3.1 中国黄牛的Y染色体类型多态性
根据染色体形态与类型,将中国地方黄牛Y染色体分为三大类:①北方黄牛均为中部或亚中部着丝粒Y染色体,属普通牛起源;②南方黄牛均为(近)端部着丝粒Y染色体,属瘤牛起源;③中原黄牛存在中部或亚中部着丝粒和(近)端部着丝粒Y染色体,属于普通牛与瘤牛混合起源[4]。
3.2 中国黄牛的mtDNA母系起源多样性
Lei等[7]分析了231头黄牛的线粒体DNA D-loop区遗传多样性,指出中国黄牛有普通牛和瘤牛两种血统。Lai等[13]研究了14个中国黄牛品种的84个个体,发现中国黄牛有普通牛和瘤牛两大支系,同时这两个支系可以细分成更多的小支系。
3.3 中国黄牛的Y-STR多态性
近年来,国内专家利用Y染色体特异性微卫星标记研究中国地方黄牛遗传多态性与父系起源。张志清[14]从10个 Y-STR标记中筛选出 UMN2404和INRA124两个标记,在秦川牛、南阳牛、晋南牛和鲁西牛中呈现多态,并且发现有瘤牛和普通牛两种父系起源。Cai等[11]对中国15个地方黄牛品种的310头公牛进行Y-STR多态性研究,发现2个Y-STR位点UMN2404和UMN0103具有明显的多态性,可以区分瘤牛和普通牛起源。辛亚平[12]对中国13个黄牛品种的19个Y-STR分析,发现有3个Y-STR位点INRA124、UMN2404和UMN0103可以区分瘤牛和普通牛起源。然而INRA124位点已经被证明在公牛与母牛中均可以扩增出相同大小的等位基因,表明这个位点不是Y染色体上特有的,不能用来区分普通牛和瘤牛起源[15]。
两个Y染色体微卫星位点INRA189和BM861是区分家牛中普通牛和瘤牛的经典标记[16,17]。在中国黄牛中这两个位点也表现出多态,在INRA189位点,发现3个等位基因,其大小分别为166、156和144bp,其中瘤牛的等位基因大小为156/144bp,普通牛为166/156bp;在BM861位点中,中国黄牛的3个等位基因大小分别为187、169和167bp,其中瘤牛在该位点的等位基因大小为187/167bp,普通牛为187/169bp。表明这2个Y-STR位点可以很好地区分中国黄牛中的普通牛与瘤牛父系起源。虽然这两个位点的等位基因大小和Edwards等[17]发现的不同,她们发现INRA189位点的大小在90~106bp之间,BM861位点的大小为156和158 bp;但是与Liu等[18]研究的INRA189和BM861的结果相似,这是因为研究的方法不同引起的。
综合前人的研究,本研究发现北方牛群体中普通牛单倍型频率较高,瘤牛在南方牛群体中较高;而在中原黄牛中普通牛和瘤牛同时存在。这种基因流动模式的形成可能是由历史事件、地理隔离以及气候环境差异等造成的。这再次证明,中国黄牛主要血统来源于普通牛和瘤牛,并在中原地区汇合。
4 结论
本研究发现2个Y-STR标记 (INRA189和BM861)可以很好地区分中国地方黄牛中的普通牛和瘤牛起源。中国地方黄牛品种的Y染色体单倍型频率各不相同。北方牛群体中普通牛单倍型频率较高,瘤牛在南方牛群体中较高,在中原黄牛中普通牛和瘤牛同时存在。
[1] 张润锋,李晓锋,陈 宏.中国地方黄牛的Y染色体遗传多样性及其进化起源[J].基因组学与应用生物学,2009,28(5):1032-1038.
[2] Ajmone-Marsan P,Garcia J F&Lenstra J A.On the origin of cattle:How aurochs became cattle and colonized the world[J].Evol.Anthropol.,2010,19:148-157.
[3] 邱怀主编.中国牛品种志[M].上海:上海科学技术出版社,1986.
[4] 陈 宏,邱 怀,詹铁生,等.中国四品种黄牛性染色体多态性的研究[J].遗传,1993,15(4):14-17.
[5] 陈幼春,曹红鹤.中国黄牛品种多样性及其保护[J].生物多样性,2001,9(3):275-283.
[6] 雷初朝,陈 宏,杨公社,等.中国部分黄牛品种mtDNA遗传多态性分析[J].遗传学报,2004,31(1):57-62.
[7] Lei C Z,Chen H,Zhang H C,et al.Origin and phylogeographical structure of Chinese cattle[J].Anim.Genet.,2006,37:579-582.
[8] Stranzinger G F,Steiger D,Kneubuhler J,et al.Y chromosome polymorphism in various breeds of cattle(Bos taurus)in Switzerland.Appl.Genet.,2007,48(3):241-245.
[9] Edwards C J,Gaillard C,Bradley D G,et al.Y-specific microsatellite polymorphisms in a range of bovid species[J].Anim.Genet.,2000,31(2):127-130.
[10] Li M H,Zerabruk M,Vangen O,et al.Reduced genetic structure of north ethiopian cattle revealed by Y-chromosome analysis[J].Heredity,2007,98:214-221.
[11] Cai X,Chen H,Wang S,et al.Polymorphisms of two Y chro-mosome microsatellites in Chinese cattle[J].Genet.Sel.Evol.,2006,38:525-534.
[12] 辛亚平.中国部分黄牛群体Y染色体微卫星多态性与分子进化及生产性能关系初步研究[D].陕西杨凌:西北农林科技大学.2007
[13] Lai S J,Liu Y P,Liu Y X,et al.Genetic diversity and origin of Chinese cattle revealed by mtDNA D-loop sequence variation[J].Mol.Phylogenet.Evol.,2006,38:146-154.
[14] 张志清.中国四个黄牛品种的父系和母系起源研究[D].陕西杨凌:西北农林科技大学.2005.
[15] Perez-Pardal L,Royo L J,Ponce De Leon F A,et al.Female segregation patterns of the putative Y-chromosome-specific microsatellite markers INRA124and INRA126do not support their use for cattle population studies[J].Anim.Genet.,2009,40:560-564.
[16] Ginja C,Penedo MCT,Melucci L,et al.Origins and genetic diversity of New World Creole cattle:inferences from mitochondrial and Y chromosome polymorphisms[J].Anim Genet,2010,41:128-141.
[17] Edwards C J,Ginja C,Kantanen J,et al.Dual origins of dairy cattle farming-evidence from a comprehensive survey of European Y-chromosomal variation[J].PLOS ONE,2011,1(6):e15922.
[18] Liu W S,Beattie CW & Ponce De Leon F A.Bovine Y chromosome microsatellite polymorphisms[J].Cytogenet.Genome.Res.,2003,102(1~4):53-58.
猜你喜欢
川北医学院学报(2022年6期)2022-06-24
小学科学(学生版)(2021年12期)2021-12-31
智慧健康(2021年17期)2021-07-30
小学科学(学生版)(2020年7期)2020-07-28
中国产前诊断杂志(电子版)(2020年1期)2020-05-21
遵义医科大学学报(2020年6期)2020-02-05
创新作文(3-4年级)(2019年3期)2019-09-03
中国漫画(2017年10期)2018-01-25
中国漫画(2017年4期)2017-06-30
创业家(2015年2期)2015-02-27
