
不同种质翅果油树表型及抗旱性比较研究
2012-07-31 03:30:48宫慧芳
浙江农业科学 2012年3期
宫慧芳,陈 惠
(山西师范大学 生命科学学院,山西 临汾 041004)
翅果油树(Elaeagnus mollis Diels.)属胡颓子科、胡颓子属的木本油料植物,目前被列为国家重点保护野生植物名录,属国家二级保护植物。自然分布狭窄仅限于山西南部的五县和陕西的户县[1-2]。翅果油树为早春蜜源植物,种仁中含有较高含量的不饱和脂肪酸亚油酸和高含量的维生素E,且出油率高、油质好,营养价值丰富[3];其根系发达,根部具根瘤菌,可改良土壤、增加土壤养分[4],因而具有较高的经济价值和生态效益,开发和利用前景广阔。
前人根据翅果油树果形分为3个生态类型,即长果型、大宫灯和小宫灯。然而,近年来山西琪尔康生物公司每年秋季从山西南部5县收集翅果油树果实,根据中果皮颜色、果实大小、形状等的差异共鉴定出222份资源品种,陈惠等[5]对其中产地为翼城的2个翅果油树种质的果实表型特征以及幼苗生长发育进行过初步研究,关于其他产地不同种质翅果油树果实表型特征、幼苗表型差异及种质间抗旱生理特性的比较研究还鲜见报道。
本研究选取产地为乡宁的果实形态不同的3个翅果油树种质,除对其果实和幼苗的表型进行观察比较外,还对干旱胁迫下3个种质的叶片保水力、质膜透性、叶绿素含量、超氧化物歧化酶(SOD)、过氧化物酶(POD)、游离脯氨酸进行分析比较。对了解翅果油树种质之间的表型差异,幼苗早期的品种分类、鉴定、选育、栽培留种有一定的借鉴。
1 材料与方法
1.1 材料
2009年9月在山西省乡宁县采集了3个翅果油树种质的果实,去掉最外层的膜质果皮,剩下带有中果皮的果实常温下保存备用。2010年5月20日,在山西师范大学生命学院温室进行种子萌发实验,6月20日3个种质分别选取生长一致的幼苗在组织培养实验室进行生理指标的测定。
1.2 播种与处理
将采集的3个翅果油树种质的果实分别人工去除木质化的中果皮和胚部的革质种皮,露胚胚部朝下插入6 cm×6 cm的营养钵中(栽培基质:花卉营养土与蛭石1∶3)每钵4粒种子,播种的深度为种子与土壤表面平齐,放置在23~25℃的露天环境,以后每2 d向浅盘中加水1次,保持基质表面湿润。待幼苗生长15 d之后采用盆栽控水法进行干旱处理。实验分3个水分胁迫梯度处理,即正常浇水(2 d浇水1次)、轻度干旱胁迫(5 d浇水1次)和重度干旱胁迫(7 d浇水1次),干旱处理2周,每个处理设3组重复,共54钵。取翅果油树自下而上第2片叶子,作为供试材料进行生理指标的测定。
1.3 相关指标测定
每个种质随机选取30个果实在电子天平上称其重量(精度为0.001 g),重复3次;用游标卡尺对3个种质的果实进行长度、宽度测量。
幼苗叶片保水力和相对电导率的测定参照马引利[6]的方法;叶绿素含量测定采用张志良等[7]的分光光度法;SOD的测定采用氮蓝四唑(NBT)法[8];POD 酶活性的测定根据 Jasdanwala 等[9]方法;脯氨酸含量的测定采用磺基水杨酸法[10]。
1.4 数据处理
实验数据用SPSS13.0软件进行统计分析。
2 结果与分析
2.1 不同种质果实、幼苗表型的观察比较
2.1.1 不同种质翅果油树果实形态观察
根据带有中果皮果实的大小、形状、果皮上沟和脊的花纹颜色等的表型观察,将本实验选取的3个供试种质表型特征(图1)描述如下。
种质1:小宫灯型,果实颗粒小,呈灯笼形,沟和脊的分界明显,沟的颜色比脊的颜色浅,沟的颜色呈灰白色,脊的颜色呈灰色。
种质2:大宫灯型,果实颗粒较大,呈灯笼形,沟和脊的分界明显,脊的颜色比沟的颜色要浅,沟的颜色呈深棕色,脊的颜色呈红棕色。
种质3:长果型,果实的颗粒大,呈梭形,沟和脊的分界明显,脊的颜色比沟的颜色深,脊的颜色呈棕色,沟的颜色呈灰白色。

图1 3个翅果油树种质的果实形态
2.1.2 不同种质翅果油树果实大小、质量的比较
种子的重量是衡量种子活力的重要指标之一,粒大、饱满的种子、内部贮藏的营养物质多,有利于种子萌发和幼苗的生长。由图1和表1可以看出,3个种质在质量、长度和宽度上有极显著差异。种质3的质量和长度要明显大于种质1和2,种质2的宽度要大于种质1和3。

表1 3个种质果实大小、质量的比较
2.1.3 不同资源种质间幼苗的生长发育和表型差异
幼苗生长的第18天,种质1的叶片没有完全展开,为淡绿色,株高为0.7~1.1 cm;种质2的叶片狭长,呈现黄绿色,株高为1.3~1.8 cm;种质3的叶片肥厚,深绿色,株高为2.0~2.2 cm。幼苗生长的第26天,种质3的生长情况明显要比种质1和2良好(图2)。
观察3个种质表型有明显差异,3个种质的叶片面积不断增大,株高增加,测量各种质株高分别为2.2~2.8 cm,2.3~3.0 cm和3.2~3.5 cm。在幼苗生长的第29天,观察到3个资源种质的株高明显增加,叶片面积明显增大,种质1叶片呈现长椭圆形,种质2叶片为卵圆形,种质3叶片呈现长卵圆形。种质3的叶面积明显大于种质1和种质2,幼苗叶片表型差异更加明显。测量各种质的株高分别为 3.0~3.3 cm,3.2~3.5 cm和 3.7~4.2 cm。

图2 翅果油树3个种质26 d苗龄的幼苗表型
2.2 不同翅果油树种质幼苗抗旱性比较
2.2.1 干旱胁迫对离体叶片保水力的影响
叶片保水力指叶片在离体条件下,保持原有水分的能力。保水力的高低与植物遗传特性有关,与细胞特性,特别是与原生质胶体特性有关,反应干旱条件下叶片抗脱水的能力,可以作为衡量植物抗旱品种筛选的简易指标[6,11]。本实验测定了轻度胁迫下离体叶片的保水力,发现不同种质叶片间叶片保水力差异极显著,种质3的叶片保水力高于种质1和2为86%,表明种质3有较强的抗脱水能力,种质2次之,种质1最差(图 3)。

图3 干旱胁迫下叶片保水力的比较
2.2.2 干旱胁迫对叶片叶绿素含量的的影响
15日苗龄的3个翅果油树种质分别进行正常浇水、轻度干旱和重度干旱处理15 d后,各取同一叶位的叶片进行叶绿素含量的测定(图4)。
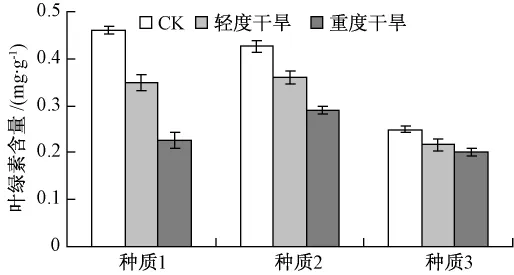
图4 干旱胁迫下叶绿素含量的变化
图4 结果表明,随着干旱程度的加重,各种质叶绿素含量均呈现下降的趋势,但不同种质间下降的幅度各有差异。种质1下降的幅度最大,种质2次之,种质3下降的幅度最小。说明3个翅果油树种质的抗旱性强弱为种质3>种质2>种质1。
2.2.3 干旱胁迫对叶片质膜透性的影响
由图5可知,随着干旱程度的加重,3个供试种质的相对电导率均呈现不同程度的上升,在重度干旱胁迫处理时,种质1,2和3的相对电导率比对照分别增加了56.63%,43.12%,27.39%。因此在干旱胁迫下,种质3的相对电导率变化较小,说明干旱胁迫对种质3的伤害最小,其抗旱能力要大于种质1和种质2。
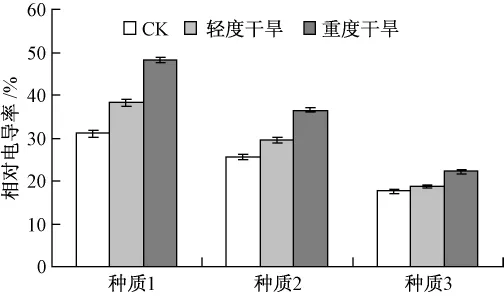
图5 干旱胁迫对细胞膜透性的影响
2.2.4 干旱胁迫对叶片保护酶活性的影响
由图6和图7可知,在干旱胁迫条件下,3个种质的SOD、POD活性均呈上升趋势,但上升的幅度有所差异。种质3的 SOD、POD活性迅速升高,且含量明显高于种质1和种质2。由此可知,种质3具有明显的清除体内活性氧的能力,而种质1和种质2抵御活性氧的能力较弱。这说明在干旱胁迫下,种质3有强的自我修复能力和生理调节能力,对逆境的抗旱能力较强。

图6 干旱胁迫下POD活性的比较
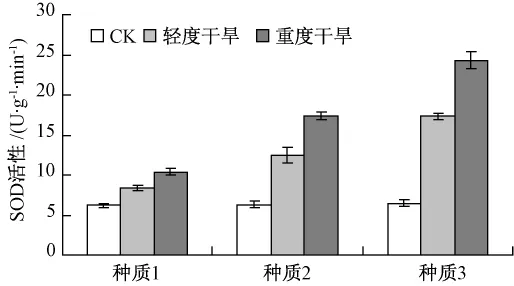
图7 干旱胁迫下SOD活性的比较
2.2.5 干旱胁迫对叶片脯氨酸含量的影响
从图8可知,在干旱胁迫下,各种质叶片脯氨酸含量均有不同程度的增长,不同种质间差异极显著,与POD和SOD酶活性变化趋势有一定的相似性。在重度干旱处理时,种质1,2和3的脯氨酸含量比对照分别增加了0.527,1.277和2.201倍。种质3叶片内脯氨酸含量增幅最大,则说明种质3在干旱胁迫下渗透调节能力最强,抗旱能力也最强。

图8 干旱胁迫下脯氨酸含量的变化
3 小结和讨论
3.1 不同翅果油树种质之间果实与幼苗之间的表型差异
选取乡宁县3个翅果油树种质为材料,研究不同翅果油树种质资源品种之间的果实、幼苗的表型差异和形态的多样性及不同种质间的抗旱生理特性差异,结果表明,3个种质在果型、坚果花纹、干果的大小、重量以及幼苗生长发育表型等方面有很大差异,而且各种质幼苗叶片面积大小与果实重量大小之间存在相关性,果型与幼苗叶形之间也有一定的相关性,与陈惠等[5]用翼城产地的2个翅果油树种质为材料研究的结果是一致的。因此,中果皮花纹、颜色、干果大小等可作为翅果油树品种鉴定的可靠指标。
3.2 不同翅果油树种质之间的抗旱生理特性差异
水分胁迫是各种环境胁迫中最为普遍的逆境因子之一。植物对干旱胁迫的响应包含着形态结构和一系列复杂的生理生化变化,并形成了植物各自遗传性机制的适应能力[12-13]。为了研究不同翅果油树种质对干旱胁迫的抵抗能力,本研究对3个种质进行了正常浇水、轻度干旱胁迫和重度干旱胁迫3种梯度处理,通过测定其离体叶片的保水力、叶绿素含量、质膜透性、POD、SOD保护酶活性和脯氨酸含量,对其抗旱性的差异进行了比较。
任庆成等研究显示:在相同的水分胁迫条件下,抗旱性强的烤烟品种叶片相对含水量下降速度较慢且叶片保水能力强[11]。本研究采用的3个翅果油树种质表现出相似的规律,种质3的叶片保水能力最强,抗旱能力最高,可以有效缓解干旱对植物细胞结构的破坏,维持较长时间的正常生命活动。
干旱胁迫引起植物光合能力下降,可能由于干旱使植物叶绿体中的光合作用片层结构受损,光合磷酸化和电子传递受阻[14]。在重度胁迫下,不同种质的翅果油树叶片叶绿素含量均有下降,但是幼苗仍可以维持正常的生命活动,这说明翅果油树具有一定的抗旱性,与刘仁涛等对翅果油树抗旱性研究的结果一致[15]。但3个种质之间存在差异,种质3叶绿素含量下降的幅度最小最抗旱,种质2次之。
保护酶系统的POD、SOD、CAT是植物细胞抵御清除活性氧,降低膜脂过氧化水平重要的保护系统[16]。在干旱胁迫下,各种质翅果油树叶片的SOD和POD酶活性均有增高,表明其自身的保护能力加强,翅果油树具有一定的抗旱潜能,也说明在本研究设计的干旱胁迫梯度下,并未对翅果油树幼苗产生严重伤害。但各种质保护酶增加的程度不同,种质3增加最多,因此抵御干旱的能力最强最为抗旱,种质2次之,种质1最弱。脯氨酸是一种渗透调节物质,可以降低因渗透失水造成的对细胞膜、酶及蛋白质结构与功能的伤害,在提高植物对各种胁迫因子抗性等方面起着十分重要的作用[17]。在干旱胁迫下,种质3幼苗叶片内的脯氨酸含量迅速增加且增加幅度最大,因此干旱适应性最强。
综上所述,根据多种生理指标的综合分析可以确定3个翅果油树种质的抗旱性顺序依次为:种质3、种质2、种质1。本研究再次证明翅果油树在形态、生理生化特性等方面具有丰富的遗传多样性,为新优品种的选育,优良种质资源的发现等方面提供了一定的方法,具有一定的参考价值。
[1]宋朝枢,徐荣章,张清华.中国珍稀濒危保护植物[M].北京:中国林业出版社,1989.
[2]贾良智,周俊.中国油脂植物[M].北京:科学出版社,1987.
[3]冯宝英,杨坪荣.翅果油树种仁化学成分分析研究[J].山西林业科技,1989(4):4-9.
[4]王毅岩.一种新的共生固氮植物:翅果油树[J].植物学报,1981,23(5):389-391.
[5]陈惠,卢英梅,申峻沛.不同栽培基质对翅果油树种子萌发的影响及不同种质幼苗生长发育的表型研究[J].山西师范大学学报:自然科学版,2010,24(3):63-66.
[6]马引利,杨纯,闫桂琴,等.土壤水分对翅果油树幼苗生理生化特性的影响[J].西北植物学报,2008,28(7):1397-1403.
[7]张志良.植物生理学实验指导[M].北京:高等教育出版社,2003.
[8]GiannopolitisC N,RiesS K.Superoxide dismutase I:Occurrence in higher plants[J].Plant Physiolgy,1977,59:309-314.
[9]Jasdanwala R T,Singh Y D,Chiony J J.Auxin metabolism in developing cotton hairs[J].Journal of Experimental Botany,1997,28:1111-1116.
[10]李霞,阎秀峰,于涛.水分胁迫对黄檗幼苗保护酶活性及脂质过氧化作用的影响[J].应用生态学报,2005,16(12):2353-2356.
[11]任庆成,陈秀华,张生杰,等.不同烤烟品种抗旱生理特征比较研究[J].西北植物学报,2009,29(10):2019-2025.
[12]Media BV.Regulation of gene expression during water deficit stress[J].Plant Growth Regulation,1999,29:22-33.
[13]Egert M,Tevini M.Influence of drought on some physiological parameters symptomatic for oxidative stress in leaves of chives(Allium schoenoprasum)[J].Envrionmental and Experimental Botany,2002,48(1):43-49.
[14]蒋龙,尹俊,孙振中.4种画眉草抗旱性比较[J].草业科学,2009,11(26):66-70.
[15]刘任涛,毕润成,任佳.翅果油树幼苗抗旱性[J].生态学杂志,2006,25(12):1528-1531.
[16]Scandalios J G.Oxygen stress and superoxide dismutase[J].Plant Physiol,1993,101(1):7 - 12.
[17]胡化广,刘建秀,周志芳,等.结缕草属植物抗旱生理机理[J].草地学报,2008,16(2):143-144.
猜你喜欢
水利水电快报(2023年2期)2023-02-21 02:34:54
今日农业(2021年14期)2021-11-25 23:57:29
今日农业(2020年13期)2020-12-15 09:08:51
中国油脂(2020年9期)2020-09-16 03:15:14
中国油脂(2019年7期)2019-08-22 09:40:32
河南水利年鉴(2017年0期)2017-05-19 02:29:16
浙江农业学报(2017年1期)2017-05-17 06:13:36
西南农业学报(2016年4期)2016-05-17 05:42:13
山西农经(2015年7期)2015-07-10 15:00:57
塑料包装(2014年3期)2014-05-07 08:29:12
