
Claudin-7—潜在的抑癌基因
2012-06-12 06:50朱煜冰李文斌
首都医科大学学报 2012年5期
丁 磊 高 宏 朱煜冰 李文斌
(1.首都医科大学附属北京世纪坛医院肿瘤外科,北京 100038;2.首都医科大学附属北京世纪坛医院肿瘤内科,北京 100038)
Claudins家族蛋白是细胞间紧密连接蛋白的主要结构之一,其表达具有组织特异性,在维持肠上皮细胞极性和紧密连接屏障功能方面起着重要作用。为研究该家族基因Claudin-7在肠道中的功能和在肿瘤发生、发展中的作用,本研究构建Claudin-7基因敲除小鼠模型并初步分析Claudin-7为可能的大肠癌抑癌基因。
1 材料与方法
1.1 主要试剂和小鼠
小鼠饲养在SPF级动物房(美国东卡大学布罗迪医学院动物中心提供),兔抗Cox-2多抗,intergrin α2多抗购自美国Cell signaling公司,兔抗Claudin-7单抗购自日本IBL公司;鼠抗GAPDH单抗购自美国Calbiochem公司;抗兔和抗鼠辣根过氧化酶标记的二抗购自美国Promega公司。
1.2 PCR反应体系
取少许小鼠尾组织溶解在TENs缓冲液(50 mmol/L TRis,0.4 mol/L NaCl,100 mmol/L EDTA,0.5%SDS)55℃水浴5h,6 mol/L NaCl充分溶解,离心取上清加95%乙醇轻柔反复混匀溶解,离心后70%冷乙醇洗涤所得即是小鼠基因DNA;PCR引物为:2.5 kb:5'-CTGCAGAGTGAGATCCTGTCTCAAAAGTAC-3';3'-CGCATCGCCTTCTATCGCCTTCTTGACGAG-5';5.7 kb:5'-CAACTCGGGCCTGCAACTGCTG-3',3'-GCAAGCCATAGCACACGCACACCATGGGAC-5';PCR反应体系:5.7 kb片段:10×LA PCR buffer 2.5 μL,2.5 mmol/L dNTP Mix 2.5 μL,Takara LA Taq 0.2 μL,Advantage 2 PCR polymerase 0.2 μL,PCR primer各1 μL,最终反应体系25 μL;94℃变性10 min,94℃变性30s,68℃退火6 min,共35个循环,最后72℃延伸10 min;2.5 kb 片段:10 ×LA PCR buffer 2.5 μL,2.5 mmol/L dNTP Mix 2.5 μL,Takara LA Taq 0.2 μL,Advantage 2 PCR polymerase 0.2 μL,PCR primer各 1 μL,最终反应体系25 μL;94℃变性10 min,94℃变性30s,68℃退火3 min共35个循环,最后72℃延伸10 min。
1.3 苏木精-伊红(Hematoxylin-Eosin,HE)染色
肠道制作成圈状,10%甲醛浸泡,制作5 μm石蜡组织切片,二甲苯脱蜡5 min 2次,梯度乙醇脱水,苏木素浸润6 min,自来水冲洗,1%乙酸1s,1%碳酸锂1s,自来水冲洗10 min,80%乙醇2 min,酸性红1 min,乙醇醇化,最后Hemo-De再脱蜡2 min 2次,封片,光学显微镜下观察。
1.4 免疫共沉淀技术
肠道组织裂解液加入intergrin α2抗体,4℃缓慢摇晃孵育过夜;将预处理过的10 μL protein A琼脂糖珠加入到和抗体孵育过夜的细胞裂解液中4℃缓慢摇晃孵育2~4h,使抗体与protein A琼脂糖珠偶连;免疫沉淀反应后,在4℃以3 000 r/min速度离心3 min,将琼脂糖珠离心至管底,将上清小心吸去,琼脂糖珠用1 mL裂解缓冲液洗3~4次,最后加入15 μL的2×SDS上样缓冲液,沸水煮5 min,以下同蛋白质印迹技术测定Claudin-7表达。
1.5 蛋白质印迹技术
(RIPA)buffer(10%SDS,4 mol/L NaCl,10%DOC,0.5 mol/L EDTA,100 mmol/L Na Pyrophosphate,0.5mol/L NaF, 10% TX-100, 1mol/L HEPES)提取肠道组织蛋白,BCA方法测定蛋白浓度,SDS-PAGE胶行蛋白分离,凝胶转移到硝酸纤维素膜,5%脱脂牛奶封闭1 h,一抗兔抗Claudin-7、Cox-2、C-jun和C-fos 4℃孵育过夜,2.5%脱脂牛奶漂洗,辣根过氧化酶标记的二抗羊抗兔孵育常温1 h,TBS/Tween漂洗3次,每次15 min,增强化学发光法ECL发光,胶片显影、定影。
1.6 统计学方法
采用SPSS11.0软件进行统计分析,计量资料以均数±标准差表示,组间比较采用方差分析。P<0.05为差异有统计学意义。
2 结果
2.1 小鼠形态及PCR基因鉴定情况
小鼠出生后6 d形态,Claudin-7基因敲除小鼠表现为形体明显缩小,伴脱水现象(图1A),野生型小鼠体质量为(3.73±0.12)g,Claudin-7基因敲除小鼠体质量为(1.28±0.14)g;两组间比较差异有统计学意义(F=12.53,P<0.001);Claudin-7基因敲除小鼠肠道内可见明显腔内出血及肠内容物变稀(箭头所指,图1 C);基因型鉴定只扩增出5.7 kb条带者为野生型小鼠,而只有2.5 kb条带者为基因敲除小鼠(图1 B)。
2.2 HE染色结果
Claudin-7基因敲除小鼠肠道上皮细胞可见广泛脱落,细胞极性的丧失,空泡形成,而野生型小鼠肠黏膜上皮细胞形态规则(图2 A);其他组织如胃、肺、膀胱、皮肤、胸腺、肾脏组织中未见明显组织破坏(图2 B);
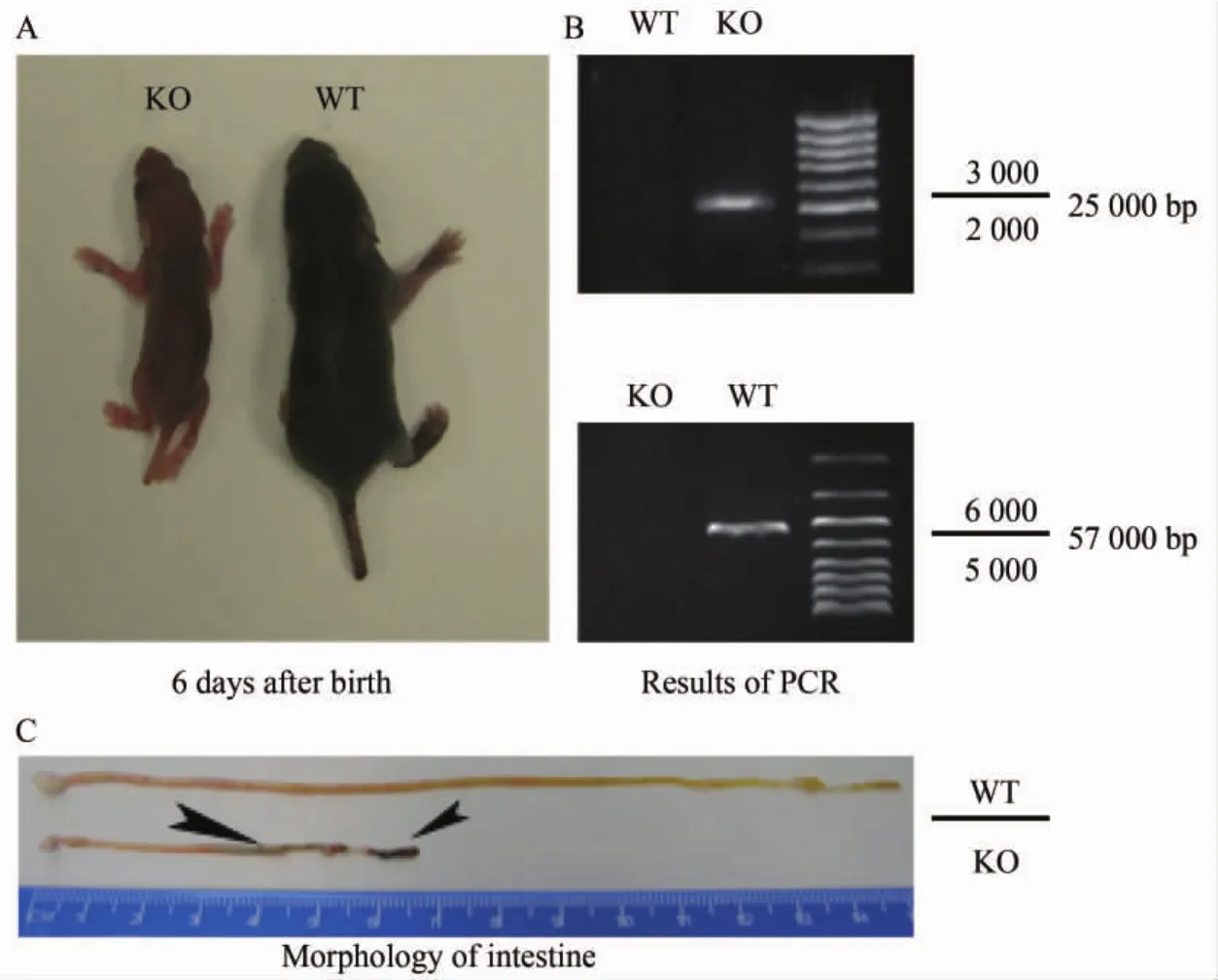
图1 Claudin-7基因敲除小鼠鉴定与形态Fig.1 Identification and morphology of Claudin-7 knockout mice
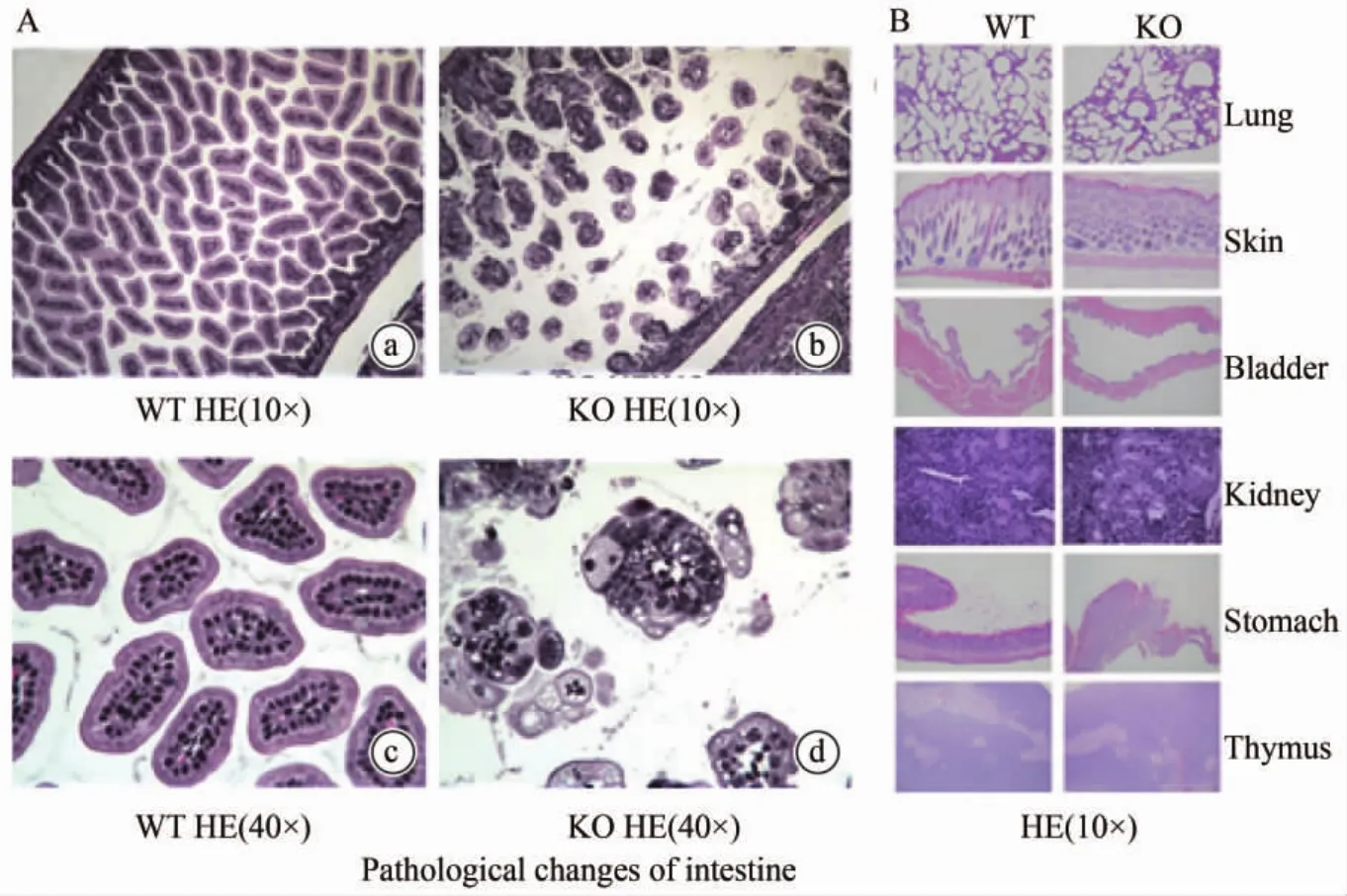
图2 Claudin-7基因敲除小鼠在不同器官病理变化(HE染色)Fig.2 Pathological changes in different organs of WT and KO mouse(HE staining).
2.3 免疫共沉淀结果
用intergrin α2抗体自肠道组织中沉淀intergrin α2蛋白后,在野生型小鼠肠道组织中可检测到Claudin-7蛋白,说明两种蛋白之间存在相互作用(图3)。
2.4 蛋白印迹结果
Claudin-7在各脏器中的表达:Claudin-7在肠道(小肠和大肠)中均匀表达(图4 A),同时在胃、肺、膀胱、皮肤、胸腺、肾脏等脏器中均有表达(图4 C),在心脏、肝脏、脑组织未见表达(图4 B)。C-jun、C-fos及Cox-2蛋白在Claudin-7基因敲除小鼠大肠和小肠中表达显著升高(P<0.001)(表1、表2,图5A)。对人类大肠癌相关组织Claudin-7蛋白检测显示Claudin-7在大肠癌组织中表达明显降低,随分化程度降低表达亦明显降低,在大肠癌肝转移组织中表达几乎丧失,蛋白表达相对值在正常大肠组织、高分化大肠癌组织、低分化大肠癌组织及大肠癌肝转移组织中分别为0.982 ±0.001、0.275 ±0.001、0.015 ±0.000、0.010 ±0.000;组间比较差异有统计学意义(F=35.4,P<0.001)(图5B)。
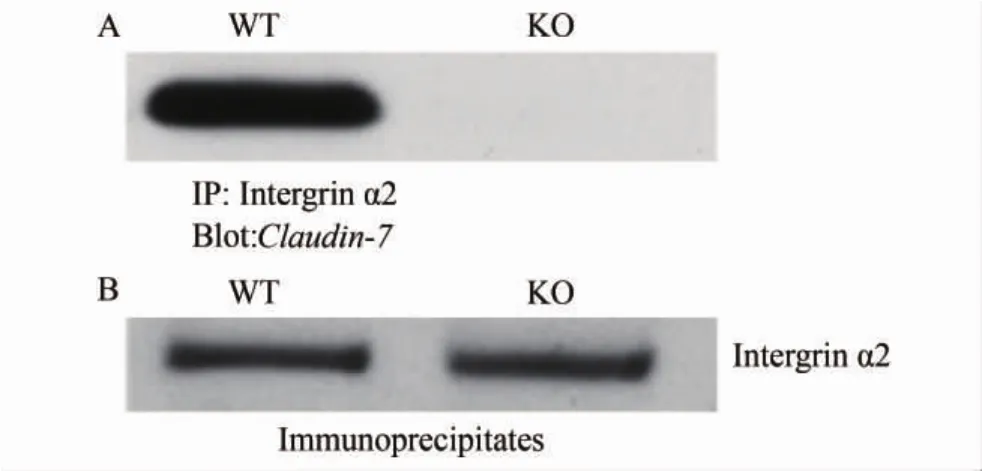
图3 免疫共沉淀结果Fig.3 Results of immunopreciptation

图4 Claudin-7在不同组织、器官表达Fig.4 Claudin-7 expression in different tissues and organs detected with Western blotting method
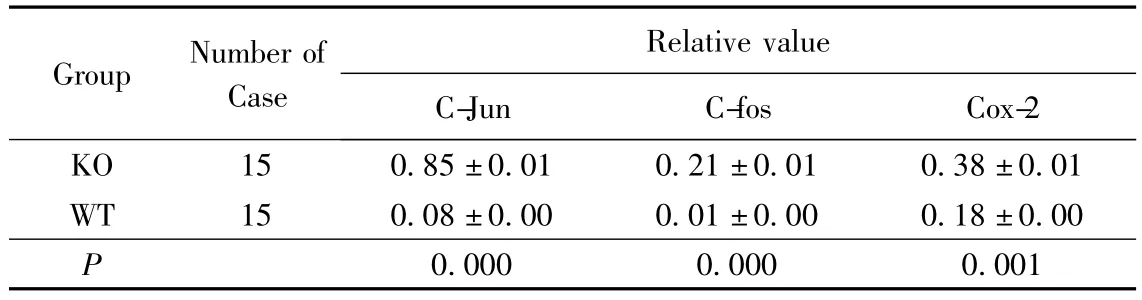
表1 C-fos、C-jun和Cox-2蛋白在Claudin-7基因敲除和野生型小鼠大肠中的表达Tab.1 Expression level of C-fos,C-jun and Cox-2 protein in large intestine of Claudin-7 gene knockout and wildtype mouse

表2 C-fos、C-jun和Cox-2蛋白在基因敲除和野生型小鼠小肠中的表达Tab.2 Expression level of C-fos,C-jun and Cox-2 protein in small intestine of Claudin-7 gene knockout and wildtype mouse
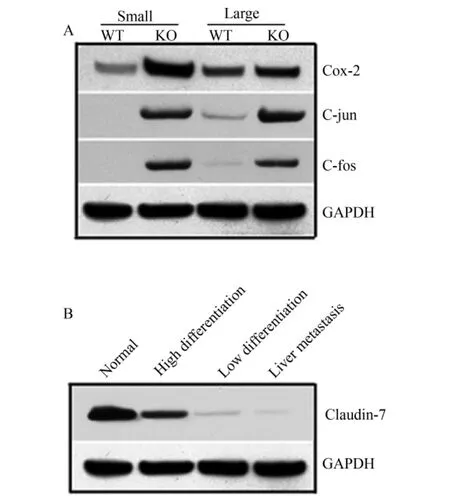
图5 Western blotting检测结果Fig.5 Results of Western blotting detection
3 讨论
紧密连接蛋白对维持正常上皮、内皮细胞的生理功能有着很重要的作用,这些蛋白质包括Occludin、Claudins家族、连接黏附分子(JAM)、PDZ表达蛋白如ZO-1、ZO-2、ZO-3 等。自从 1998 年,Furuse M[1]鉴定第一个Claudin分子以来,已有24个Claudin家族成员被识别。目前为止已有多篇关于紧密连接蛋白家族基因敲除小鼠模型的报道[2-11]:Claudin-1基因敲除小鼠表现为皮肤屏障障碍,出现明显脱水,出生后1 d死亡;Claudin-5基因敲除表现为血脑脊液屏障功能障碍,出生后当天死亡;Claudin-11基因敲除表现为血睾屏障障碍和神经系统髓鞘变性,出现耳聋;Claudin-14基因突变引起常染色体隐性耳聋;Claudin-16的基因突变会引起家族性、伴发高尿钙和肾脏钙质沉着的低镁血症,可以进行性导致慢性肾功能不全;Claudin-19基因敲除表现为雪旺细胞功能障碍;Occludin基因敲除表现为复杂的表型如出生后生长迟缓,基因敲除的雌性小鼠拒绝哺乳,雄性小鼠无生育能力,但能长久生存;ZO-1和ZO-2基因敲除表现为胚胎期死亡,ZO-3基因敲除无表型;JAM-A基因敲除表现出大肠上皮细胞通透性的改变和炎性反应的发生,但能长期生存,JAM-C完全敲除的小鼠模型体内没有成熟精子产生;Claudin-15基因敲除的小鼠出生2月后表现为近端小肠直径较正常小鼠大2倍,但能长期生存;从以上结果可以看出由于不同紧密连接蛋白的组织特异性,不同的基因突变后均出现迥异的表型,体现了各种不同紧密连接蛋白特殊的生理作用。本课题组为研究Claudin-7在机体当中的作用和在肿瘤发生、发展中潜在的功能,首次构建了Claudin-7基因敲除小鼠模型,结果发现小鼠出生时表现正常,但出生后4 d左右即停止生长,7 d左右死亡,进一步研究发现小肠和大肠黏膜表现出严重的结构破坏并伴有明显的炎性反应增生现象,显示出Claudin-7在维持肠道上皮细胞功能中有重要意义。
为进一步研究Claudin-7在机体当中的作用,本研究首先检测了各脏器Claudin-7蛋白的表达,结果发现Claudin-7在肠道中均匀表达,并且发现Claudin-7蛋白强表达且位于肠上皮细胞的顶端、侧面和基底部,不同于其他紧密连接蛋白只位于肠上皮细胞的顶端;体现了Claudin-7蛋白位置的特殊性,同时研究中还发现Claudin-7在胃、肺、肾脏、膀胱、皮肤、胸腺等脏器当中亦有表达;为此我们对表达Claudin-7的组织进行常规病理检查,结果发现只有肠道表现出显著的病理变化包括上皮细胞的严重脱落、细胞内空泡形成、极性丧失和明显的炎性反应增生现象;其他脏器除少许中性粒细胞浸润外未见明显的病理变化。结合基因敲除小鼠在出生后的生长情况,考虑小鼠的死亡与肠道吸收营养障碍有密切关系,同时表明Claudin-7并未影响小鼠的胚胎发育,很有可能是该基因敲除后影响到相关蛋白的表达或功能从而导致上皮细胞严重的免疫炎性反应,导致细胞的形态、功能变化。
为进一步探讨Claudin-7在机体中发挥作用的机制,本研究应用MicroArray初步筛选发现Claudin-7与基底膜蛋白intergrin α2存在相关性,于是采用免疫共沉淀技术进一步验证发现Claudin-7与intergrin α2蛋白免疫共沉淀,说明两者之间存在相互作用,同时免疫印迹显示C-jun、C-fos、Cox-2蛋白在Claudin-7基因敲除小鼠肠道中表达显著升高。整合素(integrin)是细胞表面重要的黏附分子受体,介导许多信号通路的发生,其中Integrin/FAK信号传导通路,在Integrin介导的肿瘤黏附转移信号转导通路中起着至关重要的作用;同时本研究发现FAK和P-FAK蛋白在Claudin-7基因敲除小鼠肠道中显著高表达,说明有可能是Claudin-7表达缺失和intergrin α2相互作用激活了下游基因FAK通路,从而激活由C-jun和C-fos形成的核转录因子AP-1(资料未发表),促进了潜在癌基因Cox-2蛋白的高表达,可能进一步诱导肠道肿瘤的发生、发展。近年来,国外诸多研究[12]表明,Cox-2参与了多种肿瘤的发生及发展,尤其与结直肠癌(CRC)等消化道肿瘤关系密切,其将成为消化道肿瘤防治研究的一个新靶点。
为进一步证明Claudin-7在大肠癌中潜在的抑癌功能,本研究应用蛋白印迹技术分别就Claudin-7蛋白在正常肠组织和不同分化大肠癌及肝转移组织中的表达进行检测,结果发现Claudin-7蛋白的表达随癌组织分化程度的降低而逐渐减弱,而在肠癌肝转移组织中表达明显降低,初步显示了Claudin-7可能在大肠癌的发生、发展方面起到潜在的抑癌作用。
Claudin-7基因敲除小鼠的建立提供了较好的小鼠肠道炎性反应性疾病模型,为研究炎性反应相关性结肠癌的发生、发展提供了良好的平台。
致谢:在此特别感谢美国东卡大学布罗迪医学院陈艳华教授给予动物模型建立支持和技术指导。
[1]Furuse M,Fujita K,Hiiragi T,et al.Claudin-1and-2:novel integral membrane proteins localizing at tight junctions with no sequence similarity to occludin[J].J Cell Biol,1998,141(7):1539-1550.
[2]Furuse M,Hata M,Furuse K,et al.Claudin-based tight junctions are crucial for the mammalian epidermal barrier:a lesson from claudin-1-deficient mice[J].J Cell Biol,2002,156(6):1099-1111.
[3]Nitta T,Hata M,Gotoh S,et al.Size-selective loosening of the blood-brain barrier in claudin-5-deficient mice[J].J Cell Biol,2003,161(3):653-660.
[4]Gow A,Southwood C M,Li J S,et al.CNS myelin and sertoli cell tight junction strands are absent in osp/claudin-11 null mice[J].Cell,1999,99(6):649- 659.
[5]Ben-Yosef T,Belyantseva I A,Saunders T L,et al.Claudin 14 knockout mice,a model for autosomal recessive deafness DFNB29,are deaf due to cochlear hair cell degeneration[J].Hum Mol Genet,2003,12(16):2049-2061.
[6]Simon D B,Lu Y,Choate K A,et al.Paracellin-1,a renal tight junction protein required for paracellular Mg2+resorption[J].Science,1999,285(5424):103-106.
[7]Miyamoto T,Morita K,Takemoto D,et al.Tight junctions in Schwann cells of peripheral myelinated axons:a lesson from claudin-19-deficient mice[J].J Cell Biol,2005,169(3):527-538.
[8]Saitou M,Furuse M,Sasaki H,et al.Complex phenotype of mice lacking occludin,a component of tight junction strands[J].Mol Biol Cell,2000,11(12):4131- 4142.
[9]Katsuno T,Umeda K,Matsui T,et al.Deficiency of zonula occluden-1 causes embryonic lethal phenotype associated with defected yolk sac angiogenesis and apoptosis of embryonic cells[J].Mol Biol Cell,2008,19(6):2465- 2475.
[10]Xu J,Kausalya P J,Phua D C,et al.Early embryonic lethality of mice lacking ZO-2,but not ZO-3,reveals critical and nonredundant roles for individual zonula occludens proteins in mammalian development[J].Mol Cell Biol,2008,28(5):1669-1678.
[11]Tamura A,Kitano Y,Hata M,et al.Megaintestine in claudin-15-deficient mice[J].Gastroenterology,2008,134(2):523-534.
[12]Oshima H,Oshima M.The inflammatory network in the gastrointestinaltumor microenvironment:lessons from mouse models[J].J Gastroenterol.2012,47(2):97-106.
猜你喜欢
当代水产(2021年10期)2022-01-12
保健医苑(2021年7期)2021-08-13
天津医科大学学报(2021年3期)2021-07-21
今日农业(2020年22期)2020-12-14
Coco薇(2016年4期)2016-04-06
中国继续医学教育(2015年4期)2016-01-07
中外医疗(2015年18期)2016-01-04
医学研究杂志(2015年9期)2015-07-01
中医研究(2013年5期)2013-03-11
河北医科大学学报(2011年12期)2011-03-25
