
基于MODIS算法的藏北高寒草甸的光能利用效率模拟
2012-06-08 08:22付刚沈振西张宪洲石培礼何永涛武建双周宇庭
草业学报 2012年1期
付刚,沈振西,张宪洲,石培礼,何永涛,武建双,周宇庭
(1.中国科学院地理科学与资源研究所 生态系统网络观测与模拟重点实验室 拉萨高原生态系统研究站,北京100101;2.中国科学院研究生院,北京100049)
光能利用效率(light use efficiency,LUE)指的是总初级生产力(gross primary production,GPP)或净初级生产力(net primary production,NPP)与植被冠层所吸收的光能量之比,它反映了植被利用光能的能力[1-5]。定量化生产力的时空变化是定量化全球碳循环的重要挑战之一,在所有的生产力模型中,LUE模型最有潜力定量化生产力的时空变化[6,7]。因此,作为光能利用效率生产力模型的重要参数,LUE的准确定量化模拟是定量化生产力时空变化和全球碳循环的基础。
自1999年12月中分辨率成像光谱仪(moderate resolution imaging spectroradiometer,MODIS)的Terra传感器成功发射以来,MODIS一直以较高的时间分辨率和空间分辨率对全球范围内的GPP和NPP进行着连续的监测[8,9]。然而,仍然有必要对MODIS生产力数据进行地表观测数据的验证[10-12]。通量观测技术是估算生态系统尺度GPP最好的方法之一,它往往被用来检验 MODIS的GPP产品精度[11-13]。研究[12,14]表明,基于 MODIS算法的LUE的低估造成了GPP的低估。因此,基于MODIS的GPP精度在很大程度上依赖于LUE的准确模拟。
高寒草甸生态系统在亚洲中部高寒环境以及世界高寒地区都极具代表性[15]。在全国范围内,高寒草甸总面积约为6.4×105km2,主要分布在青藏高原,是青藏高原典型植被类型之一[16-19]。全中国的高寒草甸生态系统共包含了约35.4Pg的碳,约相当于26.4%的中国所有草地碳库的总量[16]。此外,青藏高原高寒草甸生态系统的GPP可能会随着大气CO2浓度的增加而增加[15]。高寒草甸生态系统在中国的区域碳循环中起着举足轻重的作用[12]。因此,准确定量化模拟高寒草甸生态系统的LUE和生产力是定量化高寒草甸生态系统碳循环的重要组成部分。
Zhang等[12]对海北高寒草甸的研究表明,基于MODIS的年均GPP仅占通量观测数据估算的年均GPP的50.0%~66.7%,而LUE的低估是其主要原因。然而,利用通量观测数据对基于MODIS算法的藏北高寒草甸GPP和LUE进行验证的研究尚未见报道。因此,基于MODIS的LUE算法,本研究利用藏北高寒草甸通量观测系统的常规气象观测数据模拟了2004-2005年生长季节藏北高寒草甸生态系统的LUE,并用同期的通量观测数据进行了模型验证。
1 材料与方法
1.1 研究地概况
通量观测系统位于拉萨市当雄县草原站(91°04′E,30°30′N),该站距当雄县城约3km,地处念青唐古拉山的南缘,属丘间盆地类型,地势平坦。当雄县素有拉萨北大门之称,平均海拔4 200m,位于藏北藏南的交错地带。该地区属于高原性季风气候。降水量有明显的季节之分,80%的降水集中在生长季节的6-8月。冰冻期较长,持续3个月(11月-翌年1月)。土壤类型为高寒草甸土,土壤厚度为30~50cm。植物根系主要分布在0~20cm土层内。植被类型属于典型的高寒嵩草草甸植被,建群种主要有小嵩草(Kobresiaparva)、丝颖针茅(Stipacapillacea)、窄叶苔草(Carexmontis-everestii)等,伴生有多种密丛生嵩草。关于研究地概况的介绍也可以参考文献[20]。
1.2 野外观测数据
通量数据和气象数据取自当雄通量观测系统,该通量观测系统以开路涡度相关系统(open-path eddy covariance system)为主,并对常规气象要素进行观测。该系统自2003年7月开始运行。常规气象要素主要包括光合有效辐射(photosynthetically active radiation,mol CO2/mol光合光量子通量密度,PAR)、空气温度(air temperature,℃,Ta)、相对湿度(relative humidity,RH)、水汽压(vapor pressure,kPa)、0.05,0.10和0.50m 的土壤含水量(soil water content,m3/m3)等。
此外,在2005年生长季节进行了叶面积(leaf area index,LAI)的观测,时间间隔大约是15d,样方大小为0.5 m×0.5m,5个重复。为了得到连续的LAI数据,建立了野外观测的LAI数据和 MODIS的NDVI产品(MOD13Q1)数据的指数回归方程,其回归方程为y=0.13exp1.13x(R2=0.71;P<0.001;RMSE=0.05;y和x分别表示基于MODIS的NDVI数据和野外观测的LAI数据)。
1.3 观测的光能利用效率(LUEEC)
为了验证模拟结果,本研究依照下式计算了基于生态系统碳通量观测的光能利用效率,作为基于地表观测的光能利用效率(LUEEC),为了便于描述,在本研究中简称为观测的光能利用效率。

式中,GPP为总初级生产力,由碳通量数据依据一系列的公式[21-23]计算得到;APAR为植被吸收的光合有效辐射,由下式计算得到:

式中,PAR为光合有效辐射,取自常规气象观测系统;FPAR为植被冠层吸收的光合有效辐射比(fractional photosynthetically active radiation,FPAR),按照下式进行计算:
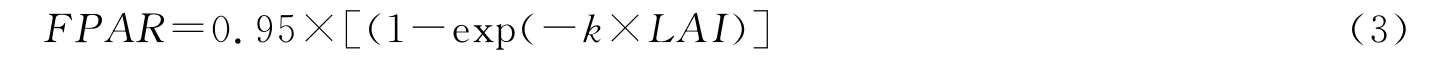
式中,k为消光系数,在本研究中,取值0.5[24];LAI为叶面积指数,由NDVI数据按照本研究前面建立的回归方程NDVI=0.13exp1.13LAI反推获得。
1.4 MODIS光能利用效率算法介绍

在MODIS光能利用效率算法中,LUE按照下式进行计算:式中,LUEmax为最大光能利用效率或潜在光能利用效率,在本研究中取0.85g C/MJ[25];Tscalar和Wscalar分别为LUEmax的温度胁迫系数和水分胁迫系数。在MODIS的LUE模型中,Tamin和VPD分别作为LUEmax的温度胁迫因子和水分胁迫因子,并分别按照式(5)和式(6)计算Tscalar和Wscalar[12,13]。

式中,Taminmax和Taminmin分别表示Tamin的最大值和最小值,在本研究中分别取MODIS算法中的默认值12.02和-8.0℃。当Tamin>12.02℃时,Tscalar值为1;当Tamin<-8.0℃时,Tscalar值为0。

式中,VPDmax和VPDmin分别表示VPD的最大值和最小值,在本研究中分别取MODIS算法中的默认值3.5和0.65kPa。当VPD>3.5kPa时,Wscalar值为0;当VPD<0.65kPa时,Wscalar值为1。
2 结果与分析
2.1 Tamin、VPD、Tscalar和Wscalar的季节变化
配对T检验表明,2年的平均Tamin(P=0.944)、VPD(P=0.566)、Tscalar(P=0.944)、Wscalar(P=0.920)都没有显著性差异。相关分析表明,2年的Tamin、VPD、Tscalar、Wscalar的相关系数分别为0.897(P<0.001),-0.372(P=0.290),0.897(P<0.001),-0.277(P=0.438)。此外,相对于水分胁迫系数而言,温度胁迫系数的值域宽度、标准差和变异系数较大(表1)。
Tamin表现出了明显的季节变化(图1):基本上呈对称的单峰曲线;2004年的峰值出现在第177~192天,而2005年的峰值比2004年的峰值出现时间晚约32d(第209~224天),但是2004年的峰值(6.76℃)要低于2005年的峰值(7.81℃);一般都在0℃以上,只在生长季初和生长季末会低于0℃。VPD曲线变化比较剧烈:基本上也呈单峰曲线,2004年的峰值(0.96kPa)出现在生长季初(第129~144天),而2005年的峰值(0.96kPa)出现在6月底7月初(第177~192天);VPD年际变动比Tamin曲线剧烈,2年的Tamin相关性达到了极显著水平,而2年的VPD相关性不显著,这从侧面反映了这一点。
Tscalar和Tamin的季节变化趋势完全一致(图2),Tscalar和Tamin2年的相关系数都为1.000(P<0.001),这从侧面反映了Tscalar和Tamin季节变化趋势的一致性。Wscalar的季节变化趋势不明显:基本上为一条恒值曲线,即几乎保持1.00不变,2004年只有第129~144,241~256和257~272天3个16d尺度的值略微低于1.00,这3个时间段的值分别为0.89,0.97和0.97;2005年也只有第145~160,161~176,177~192和241~256天4个16d尺度的值略微低于1.00,这4个时间段的值分别为0.96,0.97,0.89和0.99。
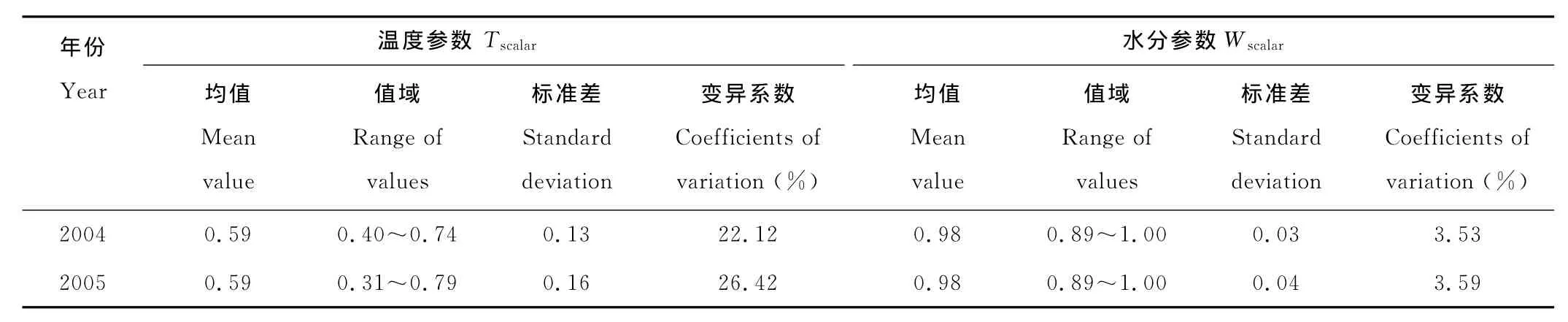
表1 光能利用效率的温度胁迫系数和水分胁迫系数的统计指标Table 1 Statistical indices temperature attenuation scalar and water attenuation scalar for light use efficiency in 2004and 2005,respectively
2.2 光能利用效率(LUEEC和LUEMODIS)的季节变化
LUEEC和LUEMODIS的季节变化趋势一致(表2,图3):LUE曲线为单峰曲线;2004年的LUEEC和LUEMODIS的峰值都出现在6月底7月初(第177~192天),其值分别为0.60和0.63g C/MJ;2005年的LUEEC的峰值(0.79 g C/MJ)出现在7月中下旬(第193~208天),而LUEMODIS的峰值(0.67g C/MJ)则出现在7月底到8月初(第209~224天),比同年的LUEEC峰值晚约16d。
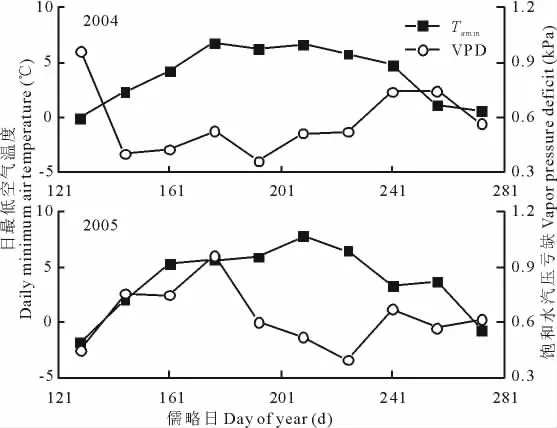
图1 日最低空气温度、饱和水汽压亏缺的季节变化Fig.1 Seasonal changes of daily minimum air temperature(Tamin)and vapor pressure deficit(VPD)
2004-2005年LUE的年际变化趋势(表2,图3)是:返青期的值都是2004年的大于2005年;生长期的LUEEC值是2004年的小于2005年,而LUEMODIS值则是2004年的大于2005年;枯黄期的值都是2004年的小于2005年;配对T检验表明,2004年的 LUEEC年均值(0.430 9±0.131 7g C/MJ)和2005年的 LUEEC年均值(0.420 6±0.159 1g C/MJ)差异不显著(P=0.814),2004年的 LUEMODIS年均值(0.495 4±0.118 3g C/MJ)和2005年的LUEMODIS年均值(0.490 3±0.129 5g C/MJ)差异也不显著(P=0.811);2004年光能利用效率的变异性小于2005年。
2.3 观测的光能利用效率(LUEEC)和 Tamin、VPD、Tscalar以及Wscalar的关系
相关分析(表3)表明,LUEEC和Tamin、Tscalar的相关性都达到了显著水平,而和VPD、Wscalar的相关性都不显著。研究[20]表明,在进行多重逐步回归分析时,截距项会影响自变量因子进入回归方程的机会。因此,本研究对有截距项和无截距项2种情况分别进行了多重逐步回归分析。回归分析结果(表4)表明,有截距项的情况下,在2004年4个自变量都没有进入回归方程;在2005年则是截距项不显著(P=0.842);无截距的情况下,在2004年和2005年都只是温度胁迫系数进入回归方程,2004年的回归方程斜率、R2、Tscalar的标准回归系数均大于2005年,而2004年的RMSE小于2005年。

图2 温度胁迫系数、水分胁迫系数的季节变化Fig.2 Seasonal changes of temperature attenuation scalar and water attenuation scalar
2.4 观测的光能利用效率(LUEEC)和模拟的光能利用效率(LUEMODIS)的比较
配对T检验表明,2004年的 LUEEC均值(0.430 9±0.131 7g C/MJ)并没有显著小于 LUEMODIS均值(0.495 4±0.118 3g C/MJ)(P=0.110),2005年的 LUEEC均值(0.420 6±0.159 1g C/MJ)也没有显著小于LUEMODIS均值(0.490 3±0.129 5g C/MJ)(P=0.099)。不同物候期的光能利用效率存在着差异:生长期和枯黄期都是LUEMODIS的值大于LUEEC的值;2005年返青期的LUEMODIS的值也大于LUEEC的值,但是2004年返青期的LUEMODIS的值小于LUEEC的值(表2)。相关分析表明,2004年的LUEEC和LUEMODIS的相关系数是0.580(P=0.039),而2005年两者的相关系数是0.673(P=0.016),这也反映了LUEEC和 LUEMODIS季节变化的一致性(图3)。
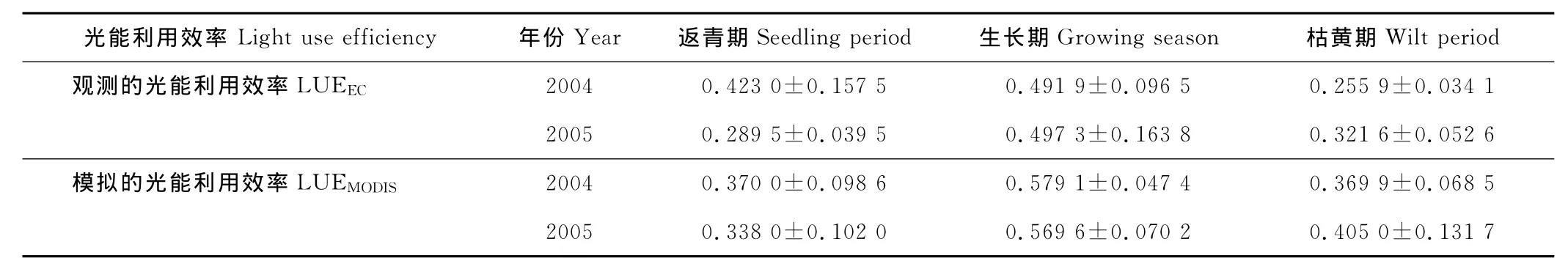
表2 不同物候期的光能利用效率Table 2 Light use efficiency of different phenophase g C/MJ
单因子线性回归分析(图4)表明,模拟的光能利用效率能解释观测的光能利用效率的季节变异;2004年和2005年回归方程的斜率相同,2004年的R2稍大于2005年,而2004年的RMSE则小于2005年,即2004年模拟的光能利用效率比2005年更多地解释了观测的光能利用效率的季节变异,同时模型误差较小,这说明2004年比2005年的模拟效果好。
为了定量化分析光能利用效率差值(LUEMODIS-LUEEC)的影响因子,本研究对光能利用效率差值和可能会造成该差值的10个因子(总初级生产力、吸收的光合有效辐射、光合有效辐射、吸收的光合有效辐射比、叶面积指数、归一化植被指数、日最低气温、饱和水汽压亏缺、温度胁迫系数以及水分胁迫系数)进行了相关分析和多重逐步回归分析(表5)。
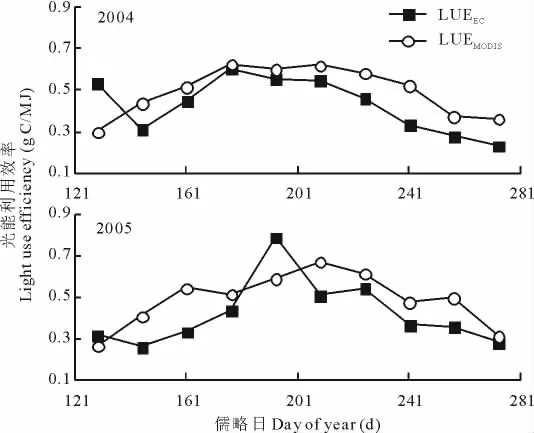
图3 光能利用效率的季节变化Fig.3 Seasonal changes of light use efficiency

表3 LUEEC与日最低气温、饱和水汽压亏缺、温度胁迫系数以及水分胁迫系数的相关分析Table 3 Correlation analysis between LUEECand daily minimum air temperature(Tamin),daytime mean vapor pressure deficit(VPD),temperature attenuation scalar(Tscalar)and water attenuation scalar(Wscalar),respectively
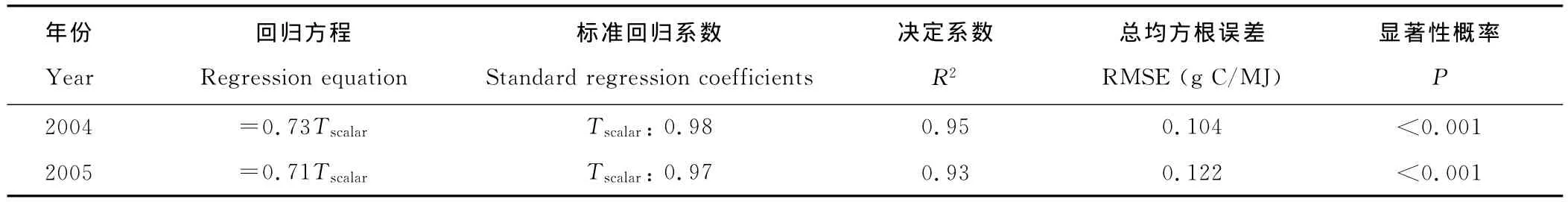
表4 LUEEC与日最低气温、饱和水汽压亏缺、温度胁迫系数以及水分胁迫系数的多重逐步回归分析Table 4 Multiple stepwise regression analysis between LUEECand daily minimum air temperature(Tamin),daytime mean vapor pressure deficit(VPD),temperature attenuation scalar(Tscalar)and water attenuation scalar(Wscalar)
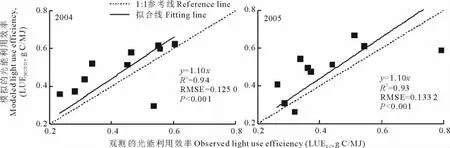
图4 模拟的光能利用效率和观测的光能利用效率的拟合Fig.4 Linear fitting between modeled light use efficiency and observed light use efficiency
相关分析表明(表5),2004年的LUEMODIS-LUEEC和吸收的光合有效辐射、吸收的光合有效辐射比、叶面积指数以及水分胁迫系数分别达到了显著或极显著相关水平,而和其他6个因子都没有达到显著相关水平;2005年的LUEMODIS-LUEEC和10个因子都没有达到显著相关水平。
多重逐步回归分析表明(表6),有截距的情况下,在2004年截距项和进入回归方程的Wscalar有着很强的共线性,在2005年则是10个参与回归的因子没有任何一个因子进入回归方程,而只有截距项;无截距的情况下,2004年只有LAI进入回归方程,并极显著地解释了37%的LUEMODIS-LUEEC季节变异,2005年则只有APAR进入了回归方程,并极显著地解释了36%的LUEMODIS-LUEEC季节变异。
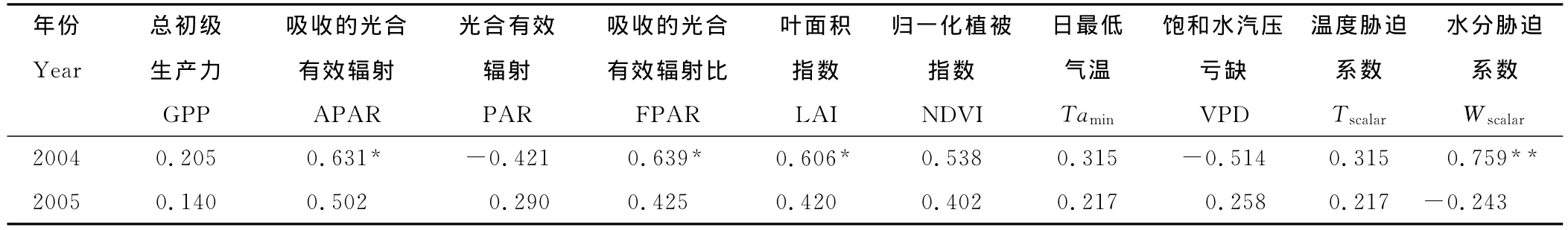
表5 LUEMODIS-LUEEC与日总初级生产力、吸收的光合有效辐射、光合有效辐射、吸收的光合有效辐射比、叶面积指数、归一化植被指数、日最低气温、饱和水汽压亏缺、温度胁迫系数以及水分胁迫系数的相关分析Table 5 Correlation analysis between LUEMODIS-LUEECand gross primary production(GPP),absorbed photosynthetically active radiation by vegetation canopy(APAR),photosynthetically active radiation(PAR),fraction photosynthetically active radiation(FPAR),leaf area index(LAI),normalized difference vegetation index(NDVI),daily minimum air temperature(Tamin),daytime mean vapor pressure deficit(VPD),temperature attenuation scalar(Tscalar)and water attenuation scalar(Wscalar),respectively

表6 LUEMODIS-LUEEC与日总初级生产力、吸收的光合有效辐射、光合有效辐射、吸收的光合有效辐射比、叶面积指数、归一化植被指数、日最低气温、饱和水汽压亏缺、温度胁迫系数以及水分胁迫系数的多重逐步回归分析Table 6 Multiple stepwise regression analysis between LUEMODIS-LUEECand gross primary production(GPP),absorbed photosynthetically active radiation by vegetation canopy(APAR),photosynthetically active radiation(PAR),fraction photosynthetically active radiation(FPAR),leaf area index(LAI),normalized difference vegetation index(NDVI),daily minimum air temperature(Tamin),daytime mean vapor pressure deficit(VPD),temperature attenuation scalar(Tscalar)and water attenuation scalar(Wscalar),respectively
3 讨论与结论
观测的光能利用效率与日最低气温的相关性强于观测的光能利用效率与饱和水汽压亏缺的相关性,且观测的光能利用效率与温度胁迫系数的相关性强于观测的光能利用效率与水分胁迫系数的相关性(表3)。观测的光能利用效率与日最低气温、饱和水汽压亏缺、温度胁迫系数以及水分胁迫系数的多重逐步回归分析结果显示,只有温度胁迫系数进入了回归方程,并极显著地解释了95%以上观测的光能利用效率的季节变异(表4)。这些结果同时表明了高寒草甸生态系统光能利用效率的季节变化主要受控于日最低空气温度。温度胁迫系数的值域宽度、标准差、变异系数分别大于水分胁迫系数的值域宽度、标准差、变异系数(表1)。配对T检验表明,温度胁迫系数均值极显著地小于水分胁迫系数均值。这些也都间接说明了温度胁迫系数比水分胁迫系数更能反应光能利用效率的季节变化。总之,相对于水分胁迫系数而言,温度胁迫系数更能够解释观测的光能利用效率的季节变异。
虽然2004年和2005年的模拟值分别高估了约14.97%和16.57%的观测值,但是模拟的光能利用效率和观测的光能利用效率差异并不显著,这表明基于MODIS算法的光能利用效率模型在定量化藏北高寒草甸生态系统光能利用效率方面具有较高的精度。
模拟的光能利用效率和观测的光能利用效率的差值的多重逐步回归分析结果显示,2004年的差异主要由LAI解释,而2005年的差异主要由APAR解释。在本研究中,LAI被用来计算FPAR,而FPAR是APAR的一个关键参数,所以,可以说模拟值和观测值之间的差异主要由APAR解释。光能利用效率差值的多重回归方程的R2值比较小,且回归方程也只是达到了显著水平,模拟光能利用效率和观测光能利用效率均值的差异性不显著在一定程度上可以解释这个结果。因此,模拟值高估的14.97%~16.57%的观测值主要受APAR的影响。
研究[6,8]表明,低温将抑制植物代谢和限制植物生长。因此,在生产力光能利用效率模型中,空气温度常作为最大光能利用效率的温度胁迫因子[6,8,26]。在气候生产力模型[18]中,空气温度也是一个非常重要的输入参数。与本研究不同的是在其他一些模型中,如植被光合模型(vegetation photosynthesis model,VPM)[26]和CASA模型[2],平均气温被用来反应温度对最大光能利用效率的胁迫作用。本研究通过相关分析比较了平均气温和日最低气温对光能利用效率的胁迫作用,结果发现,2004年和2005年平均气温和观测的光能利用效率的相关系数分别为0.515(P=0.064)和0.448(P=0.097),即都没有达到显著水平,而2年的日最低气温和观测的光能利用效率的相关性都达到了显著水平(表3);2004和2005年平均气温计算的温度胁迫系数[6,26]和观测的光能利用效率的相关系数分别为0.538(P=0.054)和0.438(P=0.103),即也都没有达到显著水平,而2年的日最低气温计算的温度胁迫系数和观测的光能利用效率的相关性都达到了显著水平(表3)。因此,在定量化藏北高寒草甸生态系统光能利用效率季节变化方面,日最低气温优于平均气温。
此外,很多研究[27-29]表明,较高的饱和水汽压亏缺将引起气孔关闭,气孔阻力增加,叶片光合速率降低,从而导致生产力降低。因此,除MODIS光能利用效率模型采用了饱和水汽压亏缺作为最大光能利用效率的水分胁迫因子外,如GLO-PEM等模型[1]也采用了饱和水汽压亏缺。
[1]Prince S D,Goward S N.Global primary production:a remote sensing approach[J].Journal of Biogeography,1995,22:815-835.
[2]Potter C S,Randerson J T,Field C B,etal.Terrestrial ecosystem production:aprocess model-based on global satellite and surface data[J].Global Biogeochemical Cycles,1993,7:811-841.
[3]王莺,夏文韬,梁天刚,等.基于MODIS植被指数的甘南草地净初级生产力时空变化研究[J].草业学报,2010,19(1):201-210.
[4]公延明,胡玉昆,阿德力麦地,等.高寒草原对气候生产力模型的适用分析[J].草业学报,2010,19(2):7-13.
[5]王雯玥,韩清芳,宗毓峥,等.不同叶型紫花苜蓿不同茬次光合效率的差异[J].草业科学,2010,27(5):50-56.
[6]Yuan W P,Liu S G,Zhou G S,etal.Deriving a light use efficiency model from eddy covariance flux data for predicting daily gross primary production across biomes[J].Agricultural and Forest Meteorology,2007,143:189-207.
[7]Wu C Y,Niu Z,Tang Q,etal.Remote estimation of gross primary production in wheat using chlorophyII-related vegetation indices[J].Agricultural and Forest Meteorology,2009,149:1015-1021.
[8]Coops N C,Jassal R S,Leuning R,etal.Incorporation of a soil water modifier into MODIS predictions of temperate Douglasfir gross primary productivity:Initial model development[J].Agricultural and Forest Meteorology,2007,147:99-109.
[9]Running S W,Nemani R R,Heinsch F A,etal.A continuous satellite-derived measure of global terrestrial primary production[J].Bioscience,2004,54:547-560.
[10]Cook B D,Bolstad P V,Naesset E,etal.Using LiDAR and quickbird data to model plant production and quantify uncertainties associated with wetland detection and land cover generalizations[J].Remote Sensing of Environment,2009,113:2366-2379.
[11]Heinsch F A,Zhao M S,Running S W,etal.Evaluation of remote sensing based terrestrial productivity from MODIS using regional tower eddy flux network observations[J].IEEE Transactions on Geoscience and Remote Sensing,2006,44:1908-1925.
[12]Zhang Y Q,Yu Q,Jiang J,etal.Calibration of Terra/MODIS gross primary production over an irrigated cropland on the North China Plain and an alpine meadow on the Tibetan Plateau[J].Global Change Biology,2008,14:757-767.
[13]Zhao M S,Heinsch F A,Nemani R R.Improvements of the MODIS terrestrial gross and primary production global data set[J].Remote Sensing of Environment,2005,95:164-176.
[14]Turner D P,Ritts W D,Cohen W B,etal.Evaluation of MODIS NPP and GPP products across multiple biomes[J].Remote Sensing of Environment,2006,(3-4):282-292.
[15]Xu L L,Zhang X Z,Shi P L,etal.Establishment of apparent quantum yield and maximum ecosystem assimilation on Tibetan Plateau alpine meadow ecosystem[J].Sciences in China Series D,2005,48:141-147.
[16]Ni J.Carbon storage in grasslands of China[J].Journal of Arid Environment,2002,50:205-218.
[17]李东,黄耀,吴琴,等.青藏高原高寒草甸生态系统土壤有机碳动态模拟研究[J].草业学报,2010,19(2):160-168.
[18]周刊社,杜军,袁雷,等.西藏怒江流域高寒草甸气候生产潜力对气候变化的响应[J].草业学报,2010,19(5):17-24.
[19]仁青吉,武高林,任国华.放牧强度对青藏高原东部高寒草甸植物群落特征的影响[J].草业学报,2009,18(5):256-261.
[20]付刚,沈振西,张宪洲,等.基于 MODIS影像的藏北高寒草甸的蒸散模拟[J].草业学报,2010,19(5):103-112.
[21]Falge E,Baldocchi D,Olson R,etal.Gap filling strategies for defensible annual sums of net ecosystem exchange[J].Agricultural and Forest Meteorology,2001,107:43-69.
[22]Fu Y,Zheng Z,Yu G,etal.Environmental influences on carbon dioxide fluxes over three grassland ecosystem in China[J].Biogeosciences,2009,6:2879-2893.
[23]Yu G R,Zhang L M,Sun X M,etal.Environmental controls over carbon exchange of three forest ecosystems in eastern China[J].Global Change Biology,2008,14:2555-2571.
[24]Ruimy A,Kergoat L,Bondeau A,etal.Comparing global models of terrestrial net primary productivity(NPP):analysis of differences in light absorption and light-use efficiency[J].Global Change Biology,1999,5:56-64.
[25]Xu L L,Zhang X Z,Shi P L,etal.Modeling the maximum apparent quantum use efficiency of alpine meadow ecosystem on Tibetan Plateau[J].Ecological Modelling,2007,208:129-134.
[26]Xiao X M,Zhang Q Y,Braswell B,etal.Modeling gross primary production of temperature deciduous broadleaf forest using satellite images and climate data[J].Remote Sensing of Environment,2004,91:256-270.
[27]Almeida A C,Landsberg J J.Evaluating methods of estimating global radiation and vapor pressure deficit using a dense network of automatic weather stations in coastal Brazil[J].Agricultural and Forest Meteorology,2003,118:237-250.
[28]同小娟,李俊,王玲.农田光能利用效率研究进展[J].生态学杂志,2008,27(6):1021-1028.
[29]同小娟,李俊,于强.农田生态系统光能利用效率及其影响因子分析[J].自然资源学报,2009,24(8):1393-1401.
猜你喜欢
成都信息工程大学学报(2022年4期)2022-11-18
青海草业(2022年2期)2022-07-23
活力(2019年21期)2019-04-01
中国新技术新产品(2019年21期)2019-01-20
今日农业(2019年14期)2019-01-04
电子技术与软件工程(2017年23期)2018-01-17
科学家(2016年12期)2017-09-29
中国市场(2017年13期)2017-06-03
河北林业科技(2016年5期)2016-11-08
植物营养与肥料学报(2012年1期)2012-10-26
