
16个树种盐胁迫下的生长表现和生理特性
2012-05-29 13:15张华新
浙江农林大学学报 2012年5期
杨 升,张华新,刘 涛
(中国林业科学研究院 林业研究所 国家林业局林木培养重点实验室,北京 100091)
土壤盐渍化是一个世界性难题。全世界大约20%的耕地和近一半的灌溉地都受到盐渍化的危害。中国盐渍土总面积约为7.5×105km2,其中沿海各省、市、自治区约1.8万km的滨海地带和岛屿沿岸,分布着各种滨海盐碱土,总面积约为5.0×106hm2[1-3]。中国沿海地区经济发达,城市化水平较高,工厂企业密集,在带动中国经济快速发展过程中有着十分重要的地位和作用[4]。随着经济和社会的发展,滨海盐碱土作为重要的土地后备资源,亟待开发、利用和保护,而最有效的方法是通过生物措施,栽植耐盐碱植物,既有经济效益,又有生态和社会效益。同时,沿海滩涂地利用,“四旁”地和城市化建设都亟需大量适应盐碱地生长的绿化植物,改善滨海地区的景观环境,增加树种多样性。目前,大量研究表明:不同类型盐胁迫条件下,植物生长和生理表现存在明显差异,植物耐盐能力表现也有所不同[5-8]。同时,滨海盐碱地以氯化钠(NaCl)为主,氯化钠胁迫下树木耐盐性研究主要集中在植物生长表现、生理生化变化及耐盐基因工程研究等方面[9-15],而针对滨海耐盐植物引种和筛选的研究较少。本研究通过对16个树种进行不同浓度盐分胁迫,比较其生长表现差异,研究耐盐性生理指标的变化规律,了解不同树种的耐盐能力,筛选和挖掘适应滨海地区的优良耐盐树种,为耐盐树种选育提供理论依据,也为滨海地区盐碱地治理提供技术支撑。
1 材料和方法
1.1 试验材料与试验设计
所用材料均为2年生苗木,部分树种是笔者于2008年5月在华北林业实验中心的温室里用种子繁殖的实生苗,种子由格林种业从美国引进。其他树种是从地方购买的实生苗或者扦插苗。总共16个树种(表1)。2009年7月,选择长势相对一致的苗木进行试验。采用完全随机区组设计,共设0,100,200,300,400,500 mmol·L-16 个盐分(NaCl)梯度。栽植苗木 10 株·梯度-1,3 次重复。装基质(草炭土(V) ∶珍珠岩(V) ∶腐殖土(V)=3 ∶1 ∶1)(5.00±0.25) kg·盆-1。施盐方式采取多次施盐,逐级递增,每次浇700 mL·盆-1,当达到预期设定的盐浓度后,浇相同量的水。盐胁迫期间,定期定量浇水,以平衡蒸发量,同时防治病虫害。花盆下垫塑料托盘,后期浇水时将流出的溶液及时倒回花盆内,以防止盐分流失。盐胁迫处理历时40 d。

表1 供试树种信息Table1 Directory of tree species for the experiment
1.2 实验方法
株高的测定:每个树种随机选取5株·处理-1,测定盐胁迫前后的株高H1和H2,株高生长量=H2-H1,株高相对生长量=盐处理的株高生长量/对照的株高生长量;盐胁迫结束后,每个树种随机选取3株·处理-1进行生物量测定,在105℃下杀青1 h,80℃烘干至恒量,称取干质量(精确到0.01g)。叶绿素[18],丙二醛(MDA)[16],超氧化物歧化酶(SOD)[16-17],脯氨酸[17]和可溶性糖[17]用 DU800 紫外可见分光光度计测定;钠离子(Na+)和氯离子(Cl-)参照王宝山等[18]和於丙军等[19]的方法,Na+用原子吸收分光光度计法测定,Cl-用自动滴定仪测定。
1.3 数据处理
用Excel 2003整理数据和绘制图表,用SPSS 13.0进行方差分析,用最小显著差(LSD)法进行多重比较。
2 结果与分析
2.1 盐胁迫对供试树种生长现象的影响
2.1.1 不同盐浓度胁迫下供试树种的盐害症状 盐胁迫过程中,10 d观察1次苗木的盐害症状,参照阎艳霞等[20]和杜中军等[21]的标准以叶片颜色变化和脱落情况进行盐害等级划分。0级:叶片颜色、生长正常;l级:部分叶尖、叶缘变黄;2级:部分叶片变黄,萎蔫,有少量叶片脱落;3级:大部分叶片变黄,萎蔫干枯,有明显落叶;4级:叶片脱落十分严重甚至死亡。对胁迫40 d的苗木受害症状列于表2。盐害症状是植物耐盐能力的直接体现。从表2可以看出:16个树种耐盐能力存在明显差异,大致可以分成3个不同层次:柽柳、唐古特白刺、西伯利亚白刺>水蜡、沙枣、药鼠李、糖朴、美国皂荚、卫矛、沙柳>杜梨、竹柳、鞑靼忍冬、醉鱼草、风箱树、美国白蜡。

表2 不同浓度盐胁迫下供试树种的受害症状Table2 Morphological changes of different tree species under salt stress (40 d)
2.1.2 不同浓度盐胁迫对株高相对生长量的影响 大部分供试树种株高相对生长量随着盐浓度的升高而呈现不同程度的下降趋势(图1),说明株高相对生长量能正面反映植物盐害的程度。在400 mmol·L-1的盐浓度下,水蜡、美国皂荚、美国白蜡和西伯利亚白刺株高相对生长量受到明显抑制,与对照相比分别降低了81.4%,66.5%,92.2%和73.4%。杜梨、沙柳和柽柳表现为低浓度促进植物高生长,在200 mmol·L-1或者 100 mmol·L-1的盐浓度下表现出最大株高相对生长量。
2.1.3 不同浓度盐胁迫对树种生物量的影响 氯化钠胁迫下,除西伯利亚白刺生物量积累随盐浓度的增加而增加外,其他各树种生物量的积累随氯化钠浓度的增加而下降,但变化规律略有不同(图2)。不同的变化规律反映了植物在盐渍环境下有机物质积累的差异。竹柳、卫矛、药鼠李、醉鱼草、美国白蜡和唐古特白刺生物量积累随盐浓度的增加而逐渐减少。低浓度(≤400 mmol·L-1)下,水蜡、鞑靼忍冬、沙枣、糖朴、风箱树和沙柳变化幅度不大,高盐浓度下生物量积累明显减少。杜梨、美国皂荚和柽柳的生物量积累随盐浓度的增加而先升后降。在400 mmol·L-1盐浓度下,竹柳、药鼠李、醉鱼草和唐古特白刺的下降程度较大,与对照相比分别下降了51.5%,44.1%,37.6%和47.3%。

图1 不同浓度盐胁迫对供试树种株高相对生长量的影响Figure1 Effect of different concentrations salt stress on relative height growth of experimental seedlings

图2 不同浓度盐胁迫对供试树种生物量的影响Figure2 Effect of different concentrations salt stress on the biomass of experimental seedlings
2.2 供试树种对盐胁迫的生理生化反应
2.2.1 盐胁迫对植物叶绿素的影响 如图3所示,大部分供试树种叶绿素质量分数随着盐浓度的增加而减少。沙柳和柽柳叶绿素随盐浓度的增加而先升后降,而唐古特白刺和西伯利亚白刺叶绿素随盐浓度的增加略有升高。在400 mmol·L-1盐胁迫下,杜梨、竹柳、鞑靼忍冬和醉鱼草的叶绿素下降幅度最大,仅为对照的52.6%,46.4%,49.4%和56.0%。在500 mmol·L-1盐胁迫下,风箱树和美国白蜡的叶绿素仅为对照的45.3%和41.1%。卫矛、水蜡、沙枣、药鼠李、糖朴、美国皂荚下降幅度次之,而沙柳、柽柳、唐古特白刺和西伯利亚白刺下降幅度很少或略有升高。

图3 不同浓度盐胁迫对供试树种叶片叶绿素的影响Figure3 Effect of different concentrations salt stress on chlorophyll contents of seedling leaves for the experiments
2.2.2 盐胁迫对丙二醛的影响 总体上,各供试树种丙二醛均随氯化钠浓度的增加而增加(图4)。鞑靼忍冬、醉鱼草和风箱树丙二醛总体明显高于其他树种,而水蜡、柽柳、唐古特白刺和西伯利亚白刺明显低于其他树种。在300 mmol·L-1盐处理水平,鞑靼忍冬和风箱树的增加量最大,与对照相比,分别增加了 57.6 nmol·g-1和 40.5 nmol·g-1。

图4 不同浓度盐胁迫对供试树种丙二醛的影响Figure4 Effect of different concentrations salt stress on MDA contents of experimental seedlings
2.2.3 盐胁迫对超氧化物歧化酶(SOD)活性的影响 盐胁迫下,各供试树种的超氧化物歧化酶活性随盐浓度的增加,变化规律有所不同。沙枣变化不明显,柽柳超氧化物歧化酶活性随盐浓度的增加而减少,水蜡、风箱树和沙柳的超氧化物歧化酶活性随盐浓度的增加而先升后降。其他树种的超氧化物歧化酶活性随盐浓度的升高而升高(图5),在300 mmol·L-1盐浓度水平,竹柳、卫矛和鞑靼忍冬超氧化物歧化酶活性与对照相比,增加幅度最大,分别为68.1%,58.5%和84.3%。
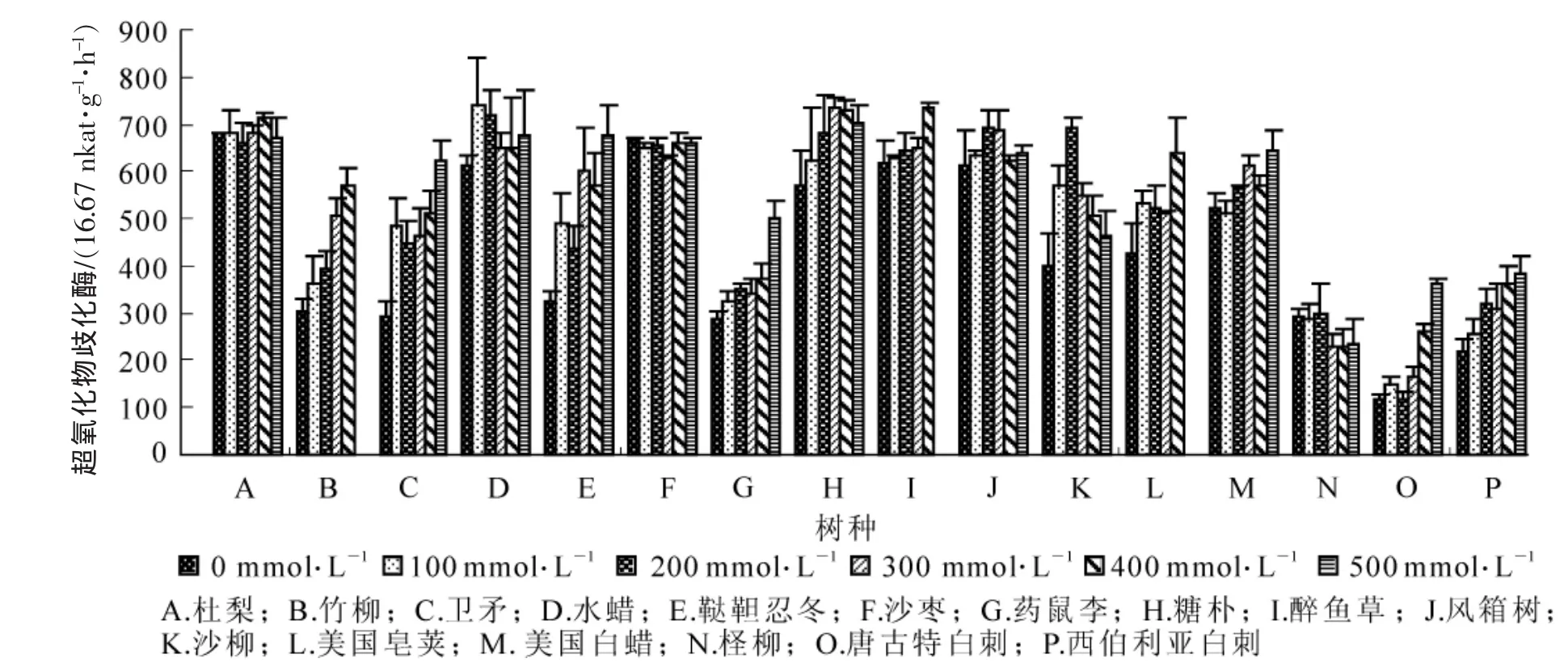
图5 不同浓度盐胁迫对供试树种超氧化物歧化酶活性的影响Figure5 Effect of different concentrations salt stress on SOD activity of experimental seedlings
2.2.4 盐胁迫对脯氨酸的影响 图6显示了不同盐浓度胁迫下各供试树种叶片中脯氨酸的变化。除沙柳变化不明显外,随盐浓度的增加,其他15个供试树种脯氨酸均有不同程度的增加。在400 mmol·L-1盐处理水平,水蜡、沙枣、糖朴和唐古特白刺与对照相比变化幅度最大,分别增加了562.8%,662.0%,184.7%和84.6%,竹柳、卫矛、药鼠李和风箱树的增加幅度最小。柽柳、唐古特白刺和西伯利亚白刺的脯氨酸总体上要高于其他树种。
图6 不同浓度盐胁迫对供试树种脯氨酸的影响Figure6 Effect of different concentrations salt stress on proline contents of experimental seedlings
2.2.5 盐胁迫对可溶性糖的影响 在盐胁迫下,各供试树种叶片中可溶性糖均有所提高。除美国皂荚的可溶性糖是随盐浓度的增加而先升后降外,其他树种可溶性糖均随盐浓度的增加而增加(图7)。在400 mmol·L-1盐浓度下,与对照相比,竹柳、卫矛、药鼠李、糖朴和醉鱼草的可溶性糖增幅最大,分别增加了72.7%,55.3%,48.5%,44.8%和68.1%,而杜梨(11.7%),鞑靼忍冬(28.1%),美国白蜡(18.4%)最小。柽柳、唐古特白刺和西伯利亚白刺可溶性糖含量总体上明显低于其他树种。
2.3 盐胁迫下供试树种矿质元素的变化

图7 不同浓度盐胁迫对供试树种可溶性糖的影响Figure7 Effect of different concentrations salt stress on soluble sugar contents of experimental seedlings

图8 不同浓度盐胁迫对供试树种根(上)、茎(中)、叶(下)内钠离子(Na+)的影响Figure8 Effect of different concentrations salt stress on Na+contents in roots(upper),stems(middle),leaves
2.3.1 盐胁迫下供试树种根、茎、叶内钠离子(Na+)变化 在各树种根、茎、叶中,钠离子(Na+)随盐浓度的增加而增加(图8)。与对照相比,400 mmol·L-1盐处理条件下,根中,竹柳、卫矛、醉鱼草和西伯利亚白刺的增幅最大,分别为275.3%,182.4%,168.4%和125.1%,而水蜡和风箱树的增幅最小,为14.5%和23.1%;茎中,竹柳(538.2%),鞑靼忍冬(649.0%),药鼠李(331.9%),醉鱼草(271.6%),美国白蜡(552.6%)的增幅最大;叶中,鞑靼忍冬、药鼠李、醉鱼草、风箱树和美国白蜡是增幅最大的,分别增加了1955.6%,1158.4%,494.8%,877.4%和362.7%。说明在高盐浓度下,根对离子的阻隔作用减弱,Na+大量流入叶片。同时,从图8中可知:在根、茎、叶3个部分中,钠离子(Na+)在16个树种中总体差异不大,但是在茎、叶中,柽柳、唐古特白刺和西伯利亚白刺的Na+要明显高于其他13个树种。这表明,柽柳、唐古特白刺和西伯利亚白刺能够大量吸收Na+,把Na+存储在叶片中,而其他树种是通过根的阻隔作用尽量减少Na+进入植物体内。
2.3.2 盐胁迫下供试树种根、茎、叶内氯离子(Cl-)变化 从图9可知:氯离子(Cl-)在各树种中总体上是随盐浓度的增加而增加,但是各树种增加幅度不同。与对照相比,400 mmol·L-1盐处理水平下,根中,增幅最大的是竹柳、卫矛、醉鱼草、沙柳和美国皂荚,依次为80.1%,117.7%,119.3%,73.9%和66.7%,水蜡、药鼠李、糖朴和美国白蜡的增幅最小;茎中,增幅最大的是竹柳、鞑靼忍冬、药鼠李、醉鱼草、风箱树和沙柳,为384.7%~154.8%;叶中,鞑靼忍冬、醉鱼草、风箱树、美国皂荚和美国白蜡增幅最大,分别为153.2%,123.0%,621.2%,203.8%和158.7%。同时可以发现,在低盐浓度下,柽柳、唐古特白刺和西伯利亚白刺茎、叶中Cl-要明显高于其他13个树种。
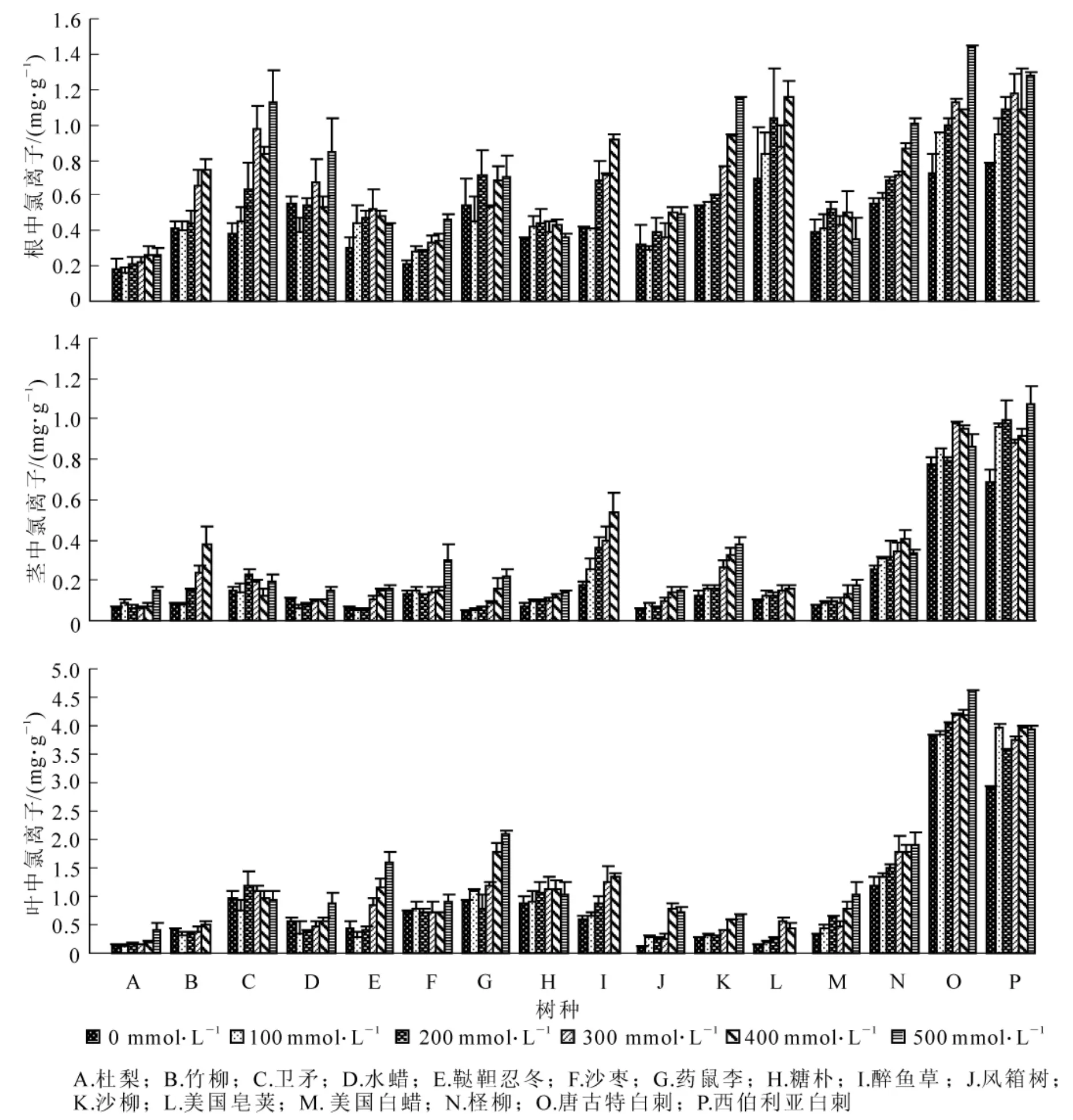
图9 不同浓度盐胁迫对供试树种根(上),茎(中),叶(下)内氯离子(Cl-)的影响Figure9 Effect of different concentrations salt stress on Cl-contents in roots (upper),stems (middle),leaves(lower) of experimental seedlings
3 讨论与结论
盐害症状表现是植物耐盐能力的直接体现。谢小丁等[22]对9种耐盐植物在黄河三角洲滨海盐碱地的耐盐能力试验中得到西伯利亚白刺的耐盐能力为1.5%,中国柽柳的耐盐能力为1.5%,沙枣的耐盐能力为1.0%,杜梨的耐盐能力为0.8%。刘昊华[23]研究显示在1.5%盐胁迫条件下,柽柳大约有50%的叶尖、叶缘受害。另外,白刺的耐盐能力强于甘肃柽柳[24]。本试验中,16个树种根据盐害症状大致可以分成3个等级:500 mmol·L-1(约土壤含盐量1.3%)盐浓度以下,可以栽植唐古特白刺、西伯利亚白刺和柽柳;卫矛、沙柳、水蜡、沙枣、药鼠李、糖朴和美国皂荚可以在400 mmol·L-1(约土壤含盐量0.8%)盐浓度以下栽植,而杜梨、竹柳、鞑靼忍冬、醉鱼草、风箱树和美国白蜡适宜栽植在300 mmol·L-1(约土壤含盐量0.5%)盐浓度以下的土壤。因此,本实验对各树种的耐盐等级划分比较准确,对于不同含盐量的滨海土壤,可以选种不同树种,做到适地适树和生物多样性。虽然树种间的耐盐能力差异是一致的,但耐盐值有所不同,可能是试验土壤结构条件和气候环境造成的,具体原因还有待进一步深入研究。
植物对盐胁迫的反应涉及到体内一系列的生理生化变化,如叶绿素、丙二醛、超氧化物歧化酶、脯氨酸等,植物的耐盐性是多种耐盐生理性状的综合表现,而各性状指标在不同植物之间存在差异。叶绿素是植物进行光合作用的重要成分,其变化直接影响生物量积累。研究报道[23-25]:在盐胁迫下,植物叶绿素随盐浓度的增加而逐渐减少。也有研究显示[11,26-27]:植物叶绿素随盐浓度增加而先升后降或者逐渐升高,并且植物表现出较强的耐盐能力。本试验结果显示:大多数树种在盐胁迫下叶绿素是减少的,说明盐渍环境影响了叶片中叶绿素的合成,也可能是加快了叶绿素的衰老和分解,而沙柳和柽柳叶绿素先升后降,唐古特白刺和西伯利亚白刺逐渐升高,说明在一定盐浓度下有利于其叶绿素的合成,它们也表现出较强耐盐能力。丙二醛是植物细胞膜脂受损程度的重要指标[17],而超氧化物歧化酶是重要的抗氧化酶,能有效清除活性氧,其活性的增加,能提高植物对膜系统的保护能力,抵抗逆境环境[28]。本试验,鞑靼忍冬和风箱树的丙二醛增加最大,说明细胞膜脂受损较重,植物耐盐能力较差,而水蜡、柽柳、唐古特白刺和西伯利亚白刺丙二醛最低,耐盐能力较强。因此,丙二醛可以作为植物耐盐性评价指标。柽柳、唐古特白刺和西伯利亚白刺的超氧化物歧化酶活性明显低于其他树种,有可能超氧化物歧化酶不是这3种植物主要的抗氧化酶,也可能是它们的细胞膜结构稳定,超氧化物歧化酶不需表现出较高活性。脯氨酸和可溶性糖是2种重要的有机渗透调节物质。Hoque等[29]研究认为:脯氨酸不仅可以作为渗透调节物质缓解盐胁迫对植物的伤害,还能起到调节细胞内微环境,有利于细胞内的各种代谢反应过程。Li等[30]认为:脯氨酸在植物胁迫过程中扮演着多重角色,它既可以作为渗透调节物质,又可以在脱水过程中保护大分子物质,同时还能够清除羟基自由基。本试验中,水蜡、沙枣、糖朴和唐古特白刺的脯氨酸增加幅度最大,竹柳、卫矛、药鼠李、糖朴和醉鱼草的可溶性糖增幅最大。说明植物都通过增加有机渗透调节物质,适应盐渍环境,但树种间选择的主要有机渗透调节物质存在差异。柽柳、唐古特白刺和西伯利亚白刺脯氨酸要明显高于其他树种,而可溶性糖则明显低于其他树种,它们的耐盐能力也高于其他树种。因此,可以初步推断脯氨酸对植物耐盐性影响较大。柽柳、唐古特白刺和西伯利亚白刺的这些生理指标与其他树种之间存在明显差异,可能与钠离子(Na+)或者氯离子(Cl-)大量进入植物叶片有关,具体原因还有待深入研究。
Na+和Cl-是造成植物盐害胁迫的主要原因,有些植物对Na+敏感,而有些对Cl-敏感[31]。本研究发现,Na+和Cl-在16个供试树种中的变化规律基本一致。此外,泌盐型植物柽柳、稀盐型植物唐古特白刺和西伯利亚白刺的Na+和C-在根中与其他树种无明显差异,而茎和叶中差异明显。根据拒盐型植物的特性,可以推断其他树种均为拒盐型植物。因此,可以根据植物根、茎、叶中的Na+或者C-差异初步推测植物的耐盐类型,从而更好地研究植物耐盐性。因此,进行植物耐盐能力和机理研究时,需要根据植物耐盐类型加以区别对待。
[1]李金耀,张富春,马纪,等.植物分子水平的耐盐机制[J].植物生理学通讯,2003,39(6):715-719.LI Jinyao,ZHANG Fuchun,MA Ji,et al.The mechanisms of salt resistance in plants in molecular levels [J].Plant Physiol J,2003,39 (6):715 - 719.
[2]王遵亲.中国盐渍土[M].北京:科学出版社,1993:325-344.
[3]徐恒刚.中国盐生植被及盐渍化生态治理[M].北京:中国农业科学技术出版社,2004:11-12.
[4]肖笃宁,解伏菊,魏建兵,等.区域生态建设与景观生态学的使命[J].应用生态学报,2004,15(10):1731-1736.XIAO Duning,XIE Fuju,WEI Jianbing,et al.Regional ecological construction and mission of landscape ecology[J].Chin J Appl Ecol,2004,15 (10):1731 - 1736.
[5]庞丙亮,曹帮华,张玉娟,等.不同盐碱胁迫对紫菀生理生化的影响[J].林业科技开发,2011,25(3):44-46.PANG Bingliang,CAO Banghua,ZHANG Yujuan,et al.Effect of saline-alkali stresses on physiological and biochemical characteristics of Aster tongolensis [J].China For Sci Technol,2011,25 (3):44 - 46.
[6]el-SAYED H E S A.Influence of NaCl and Na2SO4treatments on growth development of broad bean (Vicia faba L.)plant[J].J Life Sci,2011,5 (7):513 - 523.
[7]梅海军,王宁,李子阳,等.NaCl和Na2SO4胁迫对香樟幼苗生理特性的影响[J].西北林学院学报,2011,26(6):30 - 34.MEI Haijun,WANG Ning,LI Ziyang,et al.Effects of NaCl and Na2SO4saline stress on ecophysiological characteristics of Cinnamomum camphora seedlings [J].J Northwest For Univ,2011,26 (6):30 - 34.
[8]倪细炉,岳延峰,沈效东,等.盐胁迫对芨芨草生理响应的比较研究[J].北方园艺,2010(3):18-21.NI Xilu,YUE Yanfeng,SHEN Xiaodong,et al.Comparative studies on physiological response of Achnatherum splendens under NaCl,Na2SO4,and NaHCO3stresses [J].J Northern Gard,2010 (3):18 - 21.
[9]任丽丽,任春明,赵自国.植物耐盐性研究进展[J].山西农业科学,2010,38(5):87-90.REN Lili,REN Chunming,ZHAO Ziguo.Research advances in plant salt-tolerance [J].J Shanxi Agric Sci,2010,38(5):87 - 90.
[10]胡晓立,杨建民,陈东亮,等.NaCl胁迫对紫叶李叶片色泽的影响[J].林业科学,2010,46(12):64-69.HU Xiaoli,YANG Jianmin,CHEN Dongliang,et al.Effect of NaCl stress on leaf coloration of Prunus cerasifera var.atropurea [J].Sci Silv Sin,2010,46 (12):64 - 69.
[11]李会欣,吴明,方炎明,等.NaCl胁迫对海滨木槿叶片生理特性的影响[J].植物资源与环境学报,2010,19(3):55 - 61.LI Huixin,WU Ming,FANG Yanming,et al.Effect of NaC stress on physiological characteristics of Hibiscus hamabo leaf[J].J Plant Resour&Environ,2010,19 (3):55 - 61.
[12]姜超强,郑青松,刘兆普,等.转AtNHX1基因杨树Tr品系的耐盐性研究[J].植物生态学报,2010,34(5):563-570.JIANG Chaoqiang,ZHENG Qingsong,LIU Zhaopu,et al.Salt tolerance of transgenic poplar by the introduction of AtNHX1 gene [J].Chin J Plant Ecol,2010,34 (5):563 - 570.
[13]KAVITHA K,GEORGE S,VENKATARAMAN G,et al.A salt-inducible chloroplastic monodehydroascorbate reductase from halophyte Avicennia marina confers salt stress tolerance on transgenic plants [J].Biochimie,2010,92(10):1321 - 1329.
[14]刘昀,邓银霞,郑易之.植物耐盐的分子机理研究进展[J].安徽农业科学,2010,38(12):6087-6089.LIU Yun,DENG Yinxia,ZHENG Yizhi.Research advances on the molecular mechanism of plant salt-tolerance[J].J Anhui Agriic Sci,2010,38 (12):6087 - 6089.
[15]TANG Renjie,LIU Hua,BAO Yan,et al.The woody plant poplar has a functionally conserved salt sensitive pathway in response to salinity stress [J].Plant Mol Biol,2010,74:367 - 380.
[16]郝建军,康宗利,于洋.植物生理学实验技术[M].北京:化学工业出版社,2007.
[17]李合生,孙群,赵世杰,等.植物生理生化实验原理和技术[M].北京:高等教育出版社,2000.
[18]王宝山,赵可夫.小麦叶片中Na,K提取方法的比较[J].植物生理学通讯,1995,31(1):50-52.WANG Baoshan,ZHAO Kefu.Comparison of extract method for Na,K in leaves of wheat [J].Plant Physiol J,1995,31 (1):50 - 52.
[19]於丙军,罗庆云,曹爱忠,等.栽培大豆和野生大豆耐盐性及离子效应的比较[J].植物资源与环境学报,2001,10(1):25 - 29.YU Bingjun,LUO Qingyun,CAO Aizhong,et al.Comparison of salt tolerance and ion effect in cultivated and wild soybean [J].J Plant Resour Environ,2001,10 (1):25 - 29.
[20]阎艳霞,王玉魁,张东.不同枣品种对NaCl胁迫的适应性研究[J].河南农业大学学报,2008,42(4):338-341.YAN Yanxia,WANG Yukui,ZHANG Dong.Study on physiological adaptability of different Jujuba species to NaCl stress [J].J Henan Agric Univ,2008,42 (4):338 - 341.
[21]杜中军,翟衡,罗新书,等.苹果砧木耐盐性鉴定及其指标判定[J].果树学报,2002,19(1):4-7.DU Zhongjun,ZHAI Heng,LUO Xinshu,et al.Salt tolerance identification on apple rootstocks[J].J Fruit Sci,2002,19 (1):4 - 7.
[22]谢小丁,邵秋玲,李扬.9种耐盐植物在滨海盐碱地的耐盐能力试验[J].湖北农业科学,2007,46(4):559-561.XIE Xiaoding,SHAO Qiuling,LI Yang.Salt resistance of the 9 kinds of representative halophyte on the saline-alkaline land [J].Hubei Agric Sci,2007,46 (4):559 - 561.
[23]刘昊华,虞毅,丁国栋,等.4种滨海造林树种耐盐性评价[J].东北林业大学学报,2011,39(7):8-11.LIU Haohua,YU Yi,DING Guodong,et al.Evaluation on salt-tolerance of four coastal tree species [J].J Northeast For Univ,2011,39 (7):8 - 11.
[24]倪细炉,岳延峰,田英,等.4种盐生植物抗盐能力的综合评价[J].中国农学通报,2010,26(6):138-141.NI Xilu,YUE Yanfeng,TIAN Ying,et al.Comprehensive evaluation of salt-resistance traits in four halophytes[J].Chin Agric Sci Bull,2010,26 (6):138 - 141.
[25]邱凤英,廖宝文,肖复明.半红树植物杨叶肖槿幼苗耐盐性研究[J].林业科学研究,2011,24(1):51-55.QIU Fengying,LIAO Baowen,XIAO Fuming.Salt tolerance of semi-mangrove plant Thespesia populnea seedlings[J].For Res,2011,24 (1):51 - 55.
[26]PUSHPAM R,RANGASAMY S R S.Variations in chlorophyll contents of rice in relation to salinity [J].Crop Res,2000,20 (2):197 - 200.
[27]魏秀君,殷云龙,芦治国,等.NaCl胁迫对5种绿化植物幼苗生长和生理指标的影响及耐盐性综合评价[J].植物资源与环境学报,2011,20(2):35-42.WEI Xiujun,YIN Yunlong,LU Zhiguo,et al.Effects of NaCl stress on growth and physiological indexes of five greening plant seedlings and comprehensive evaluation of their salt tolerance [J].J Plant Resour Environ,2011,20(2):35 - 42.
[28]孙海菁,王树凤,陈益泰.盐胁迫对6个树种的生长及生理指标的影响[J].林业科学研究,2009,22(3):315-324.SUN Haijing,WANG Shufeng,CHEN Yitai.Effects of salt stress on growth and physiological index of 6 tree species[J].For Res,2009,22 (3):315 - 324.
[29]HOQUE M A,OKUMA E,BANU M N A,et al.Exogenous proline mitigates the detrimental effects of salt stress more than exogenous betaine by increasing antioxidant enzyme activities [J].J Plant Physiol,2007,164 (5):553-561.
[30]LI Gang,WAN Shuwen,ZHOU Jian,et al.Leaf chlorophyll fluoresecence,hyperspectral reflectance,pigments content,malondialdehyde and proline accumlation responses of castor bean (Ricinus communis L.) seedlings to salt stress levels [J].Indl Crops Prod,2010,31:13 - 19.
[31]赵可夫,范海.盐生植物及其对盐渍生境的适应生理[M].北京:科学出版社,2005:209-210.
猜你喜欢
草业学报(2022年5期)2022-07-08
青海农林科技(2022年1期)2022-03-05
食品与健康(2021年9期)2021-09-10
河北林业科技(2020年3期)2020-03-23
植物资源与环境学报(2019年1期)2019-04-02
农学学报(2018年10期)2018-11-01
西南农业学报(2018年9期)2018-10-19
林业与生态(2018年1期)2018-01-15
农家科技下旬刊(2017年6期)2017-07-05
作文与考试·初中版(2017年5期)2017-03-01
