
多因子对日本囊对虾Marsupenaeus japonicus氮磷代谢的影响*
2012-05-09 08:27:36孙成波李义军
中山大学学报(自然科学版)(中英文) 2012年2期
孙成波,李 婷 ,李义军,王 平
(1.广东海洋大学水产学院,广东 湛江 524025;2.广东高校热带海产无脊椎动物养殖工程技术研究中心,广东 湛江 524025;3.海南省昌江南疆生物技术有限公司,海南 三亚 572000)
日本囊对虾Marsupenaeusjaponicus,俗称花虾、车虾、斑节虾,自然分布很广,澳大利亚、红海、坦桑尼亚、印度、菲律宾、日本及我国海域均有分布。其肉质鲜嫩,营养丰富,适合盐度较高地区养殖,耐低温、耐干能力强,适合鲜活虾出售,售价高,利润空间大,一般为斑节对虾的4倍,南美白对虾的8倍,但由于其抗病性较弱,后期养殖难度大,日本囊对虾一直以来养殖成功率低,亩产低。近年来开展的日本囊对虾的研究主要集中在人工育苗及养殖方面、日本对虾营养学方面以及日本对虾病害防治技术方面[1-4],但对其能量代谢方面的研究尚未进行,为最大限度地发挥其生长潜能,提高输入能量的利用率和加强养殖系统生态的调控,开展日本囊对虾能量代谢的研究是很有必要的。
池塘养殖的对虾,其生长和代谢的能量主要来源于人工投喂的饵料,分析对虾池塘养殖系统可以看出,未被对虾利用而存留于池塘中的输入能量是造成池塘和近海水域污染的主要原因[5]。为提高养虾池塘输入能量的利用率人们正在探索综合养殖,以其多层次、多渠道利用虾池能量,国内关于这方面的研究有李松青等[6]的摄食对凡纳滨对虾氮磷排泄率的影响,宋协法等[7]的温度和盐度对凡纳滨对虾氮磷排泄率的影响,王兴强[8]的凡纳滨对虾生长和生物能量学的初步研究,刘鹏[9]的温度和盐度双因子交互作用对凡纳滨对虾生长、代谢的影响研究, 董双林等[10]的温度和体质量对日本沼虾能量收支的影响,林小涛等[11]的罗氏沼虾亲虾的能量代谢研究,张硕等[12]的温度、体质量、盐度和摄食状态对中国对虾耗氧率和排氨率的影响,吴立新等[13]的饥饿和再投喂对日本囊对虾代谢率影响的研究等等。本文从温度、体质量、盐度、pH值和摄食状态等多个环境因子对日本囊对虾氮、磷代谢的影响展开研究,探索对虾的一些重要生理生态学规律,研究虾类代谢强度的变化,为养殖生产上投饵和水质管理提供依据。
1 材料与方法
1.1 实验材料
实验用日本囊对虾M.japonicus均来源于海南省昌江南疆生物技术有限公司三联对虾养殖基地的日本囊对虾池塘,为健康活泼的个体,在设计的温度、盐度、pH值下驯养5 d,并于实验前停食2 d。实验用海水取自三联基地蓄水池。
1.2 多因子对日本囊对虾氮磷代谢的影响
1.2.1 温度和体质量 实验于5 L小桶中进行,每个小桶中各放2尾虾、1 L海水,实验温度设20、25和30 ℃ 3个梯度,体质量设3个规格,各温度体质量条件下均设3组平行,同时设置空白实验对照,实验在白天持续进行6 h,实验过程中使用恒温电热棒控制温度,实验用海水盐度为17‰,pH为8.5。实验结束后测定水中的无机磷、亚硝酸氮、硝酸氮、氨氨的含量。
1.2.2 盐度 实验于5 L小桶中进行,每个小桶中各放2尾虾、1 L海水,盐度设置10‰、17‰、24‰和31‰等4个梯度,用蒸馏水和海盐加正常海水配制不同盐度的海水。实验水温为25 ℃、pH值为8.5,对虾体质量平均为(6.739±0.023)g,每一盐度下设置3个重复,实验持续6 h。
1.2.3 pH值 实验于5 L小桶中进行,每个小桶中各放2尾虾、1 L海水, pH值设置7.5、8.0、8.5和9.0等4个梯度,用Na2CO3、NaHCO3、HCl调配不同pH值的海水。实验水温为25 ℃、盐度为17‰,对虾体质量平均为(6.749±0.01)g,每个pH值条件下设置3个重复,实验持续6 h。
1.2.4 摄食状态 饥饿2 d后的部分对虾投喂配合饲料,停止摄食后进入实验作为摄食组。实验水温为25 ℃、盐度为17‰、pH为8.5,虾体平均质量(6.729±0.028)g,摄食组与饥饿组均设置3个重复,实验持续6 h。
1.3 理化因子的测定
2 结果与分析
2.1 温度和体质量对氮磷代谢的影响
不同温度、不同体质量对日本囊对虾氮、磷的代谢率存在影响,随温度的上升,日本囊对虾磷、亚硝酸氮、硝酸氮、氨氮的代谢率均上升,经方差分析,温度对日本囊对虾氮磷代谢的影响极显著(P<0.01)。随体质量的增加,亚硝酸氮的代谢率增大,而无机磷、硝酸氮、氨氮的代谢率与体质量呈负相关,经方差分析,体质量对日本囊对虾氮磷代谢的影响亦极显著(P<0.01)。见表1。
2.2 盐度对日本囊对虾氮磷代谢的影响
随盐度的增加,日本囊对虾(6.739±0.023)g磷、亚硝酸氮、氨氮的代谢率均提高,见表2。当盐度为31‰时,磷代谢率达到(0.736±0.002)μg·g-1·h-1,亚硝酸氮代谢率为(0.0778±0.011)μg·g-1·h-1。硝酸氮、氨氮的代谢率随着盐度的增加而下降,当盐度为31‰时,硝酸氮代谢率降为(0.257±0.207)μg·g-1·h-1,氨氮代谢率降为(4.445±0.259)μg·g-1·h-1。经方差分析,盐度对磷、硝酸氮、氨氮代谢率的影响极显著(P<0.01),对亚硝酸氮代谢率的影响显著(P<0.05)。
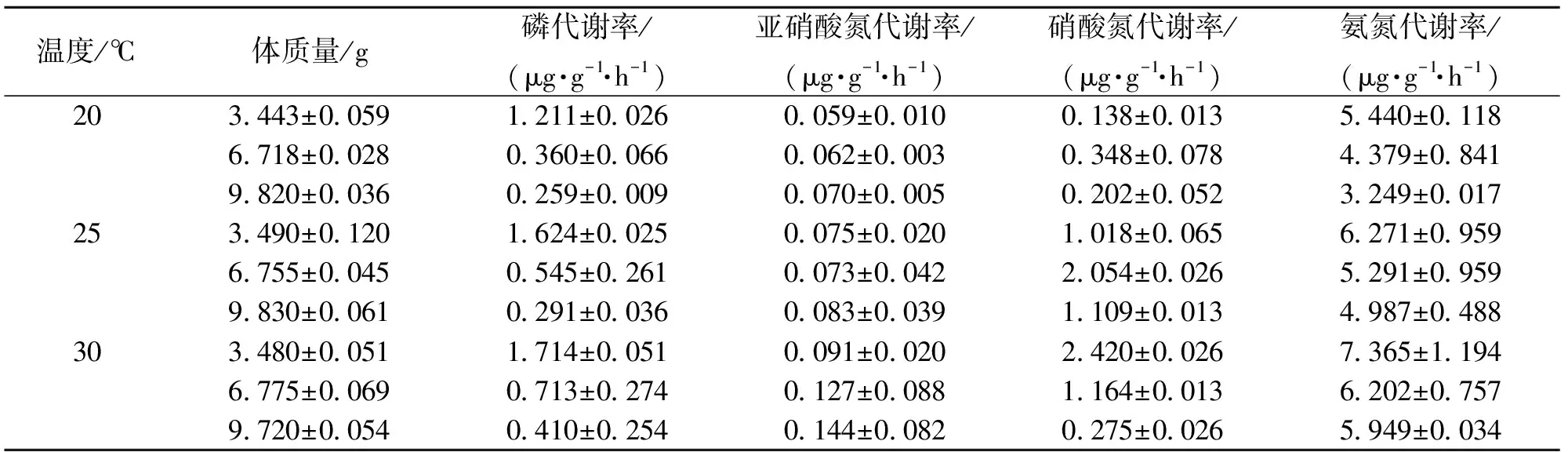
表1 不同温度下不同体质量的日本囊对虾N、P的代谢率变化

表2 不同盐度下日本囊对虾N、P的代谢率变化
2.3 pH值对日本囊对虾氮磷代谢的影响
随着pH值的增加,日本囊对虾(6.749±0.013)g磷的代谢率明显下降,pH值为9.0时,磷的代谢率最低为(1.398±0.106)μg·g-1·h-1;氨氮、硝酸氮的代谢率随着pH值的增加而升高,pH值为9.0时,氨氮、硝酸氮的代谢率分别达到(2.824±0.031)、(9.123±0.275)μg·g-1·h-1;当亚硝酸氮的代谢率在pH值为8.5时达到最大值(0.082±0.067)μg·g-1·h-1,后又呈下降趋势。经方差分析,pH值对磷、氨氮代谢率的影响极显著(P<0.01),对硝酸氮代谢率的影响显著(P<0.05),对亚硝酸氮代谢率的影响不显著(P>0.05)。
2.4 摄食状态对日本囊对虾氮磷代谢的影响
摄食状态对日本囊对虾的代谢存在明显的影响。摄食组的磷、亚硝酸氮、氨氮代谢率极显著大于饥饿组(P<0.01)。日本囊对虾(6.729±0.028)g摄食配合饲料,饱食状态下磷、亚硝酸氮、硝酸氮、氨氮的代谢率比饥饿状态下分别提高了272.02%、91.67%、795.38%、98.54%,见表4。这表明摄食时蛋白质代谢增加显著。
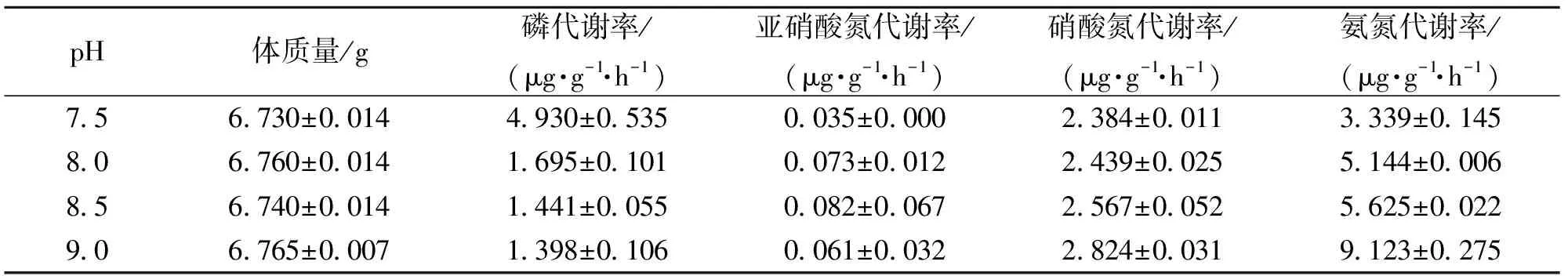
表3 不同pH下日本囊对虾N、P的代谢率变化
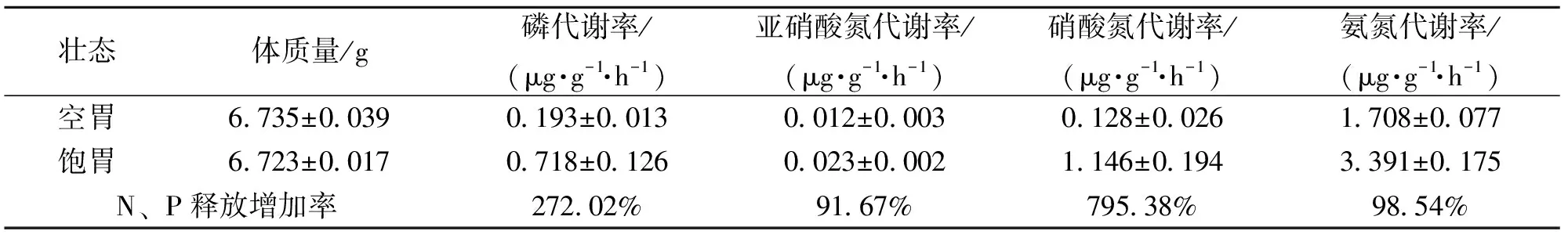
表4 不同摄食状态下日本囊对虾N、P的代谢率变化
3 讨 论
3.1 温度和体质量对日本囊对虾氮磷代谢的影响
随温度的上升,日本囊对虾磷、亚硝酸氮、硝酸氮、氨氮的代谢率均有所提高,这说明随着温度的升高,对虾同其他变温动物一样,组织器官的活动性能提高,体内的各种生化反应速度加快,致使呼吸和代谢加快,对能量的消耗增大,这与其他学者的研究结果相似[10,15-16]。随着体质量的增加,日本囊对虾磷、硝酸氮、氨氮的代谢率均极显著下降。这与林小涛等[11]在罗氏沼虾亲虾的能量代谢研究过程中,发现罗氏沼虾的NH3-N排泄率与体质量的关系呈曲线负相关的结论是相同的。
3.2 盐度对日本囊对虾氮磷代谢的影响
养殖海水盐度对海洋生物,特别是渗透调变类动物的生理状况有直接的影响。对虾与绝大多数的海洋无脊椎动物一样,属于渗透调变类(Osmoregulator)动物[17]。当外界盐度发生变化时,对虾通过改变代谢状况,籍助于离子调节和细胞内游离氨基酸库的启动或关闭,调节体内的渗透浓度,实现有机体与海水之间的水交换以及细胞和组织之间的水分平衡,以适应不同的盐度环境[18-19]。Dallavia[20]曾报道小长臂虾Dalaemoneteesantennarius在体液达到等渗点时其耗氧率最小,代谢所需要的能量最少。施正峰等[21]的实验证明了日本沼虾Macrobrachiumniponense在盐度3‰时其代谢和排泄耗能最少,有利于提高摄食能量的利用率。
本实验在盐度10‰~31‰范围内进行,随着盐度的升高无机磷、亚硝酸氮的代谢率增大,硝酸氮、氨氮的代谢率减少,这是由于日本囊对虾的最适盐度范围为18‰~30‰,当对虾处于最适盐度范围时能量消耗减少,摄入能量的利用率提高。

3.3 pH值对日本囊对虾氮磷代谢的影响
pH值即水体的酸碱度,是综合反映水中化学成分变化的一个主要指标,它影响到水中各种生物,包括各种微生物的生长繁殖,调节水体pH值在水产动物生长最理想的范围内,是水产养殖获得高产稳产的必要条件之一。虾池水正常pH值范围为7.5~8.5左右,最理想范围为8.0~8.6。当pH值过高时,虾体对于氨氮的分解代射增加,会使水中的氨氮量增加,使水体恶化,不利于虾体生长。而当pH值过低时,虾体血液中pH也会下降,血液中氧的分压降低,造成运输氧气的蛋白质功能障碍,使水产动物组织细胞内缺氧,造成虾的呼吸困难,吃食量减少,严重时引起死亡,还有会引起硫化氢含量增加。同时,pH值的高低与氮磷的排泄也存在一定的联系。由表3可知,随着pH值的增加,日本囊对虾磷的代谢率明显下降,氨氮、硝酸氮的代谢率提高,亚硝酸氮的代谢率在pH值为8.5时达到最大值,后又呈下降趋势。目前pH值对氮磷释放的作用机制的研究较少,还有待进一步的探讨研究。
3.4 摄食状态对日本囊对虾氮磷代谢的影响
摄食水平是影响虾类生长和能量收支的一个重要因素。摄食引起的代谢增加称为特殊动力作用,也称为体增热,表现为体热的增加,是在摄入的食物转化为体内可利用的能源物质过程中的能量消耗。由于特殊动力作用主要是蛋白质代谢引起的,食物中的蛋白质含量对体增热具有相当大的影响,摄食高蛋白质食物的代谢率大于摄食低蛋白质食物的代谢率。Nelson等[26]研究表明罗氏沼虾稚虾有特殊动力作用,认为摄食颤蚓后的代谢率比标准代谢率增加39.4%,摄食两种配合饲料的代谢率比标准代谢率分别增大了19.7%和7.1%。张硕等[12]认为中国对虾摄食沙蚕和配合饲料的耗氧率比饥饿状态时分别增大8.319%和2.913%,而排氨率分别增大131.8%和93.6%,明显看出排氨率的增加大于耗氧率的变化,反映出中国对虾的特殊动力作用主要是蛋白质代谢造成的,而且动物饵料的特殊动力作用大于配合饲料。
本研究发现,摄食状态日本囊对虾的氮磷代谢极显著大于饥饿组。摄食组的日本囊对虾在摄食后氮磷代谢率明显提高,这一增加的部分称外源氮、磷排泄率,外源氮、磷排泄率主要受到食物的营养组成、日粮水平和投饲方式等的影响。李松青等[6]在研究摄食对凡纳滨对虾耗氧率和氮、磷排泄率的影响时,发现凡纳滨对虾摄食后的氮排泄率平均升高40%以上。吴立新等[13]在研究饥饿和再投喂对日本囊对虾代谢率的影响时,发现日本囊对虾在饥饿时,完全靠消耗自身的蛋白质来维持生命,其代谢降低,与本实验研究结果相符。
参考文献:
[1]林琼武,单保党,黄加祺,等.北方地区日本对虾育苗业中存在的几个问题[J].中山大学学报:自然科学版,2000,39(增刊):28-33.
[2]王军霞,王维娜,王亚斌,等.日本对虾的营养需要[J].海洋通报,2003,22(5):78-85.
[3]李才文,管越强,俞志明.盐度变化对日本对虾暴发白斑综合症病毒病的影响[J].海洋环境科学,2002,2(4):6-10.
[4]田德纯.日本对虾荧光极毛杆菌病防治初探[J].中国水产,2001(2):54-55.
[5]王清印.对虾养殖与近岸水环境保护[J].国外水产,1994,33(77):1-4.
[6]李松青,林小涛,李卓佳,等.摄食对凡纳滨对虾耗氧率和氮、磷排泄率的影响[J].热带海洋学报,2006,25(2):44-48.
[7]宋协法,刘鹏,葛长字.温度、盐度交互作用对凡纳滨对虾耗氧和氨氮、磷排泄的影响[J].渔业现代化,2009,(2):24-32.
[8]王兴强.凡纳滨对虾生长和生物能量学的初步研究[D].中国海洋大学,2004:48-65.
[9]刘鹏.温度和盐度双因子交互作用对凡纳滨对虾生长、代谢的影响研究[D].青岛:中国海洋大学,2009.
[10]董双林,堵南山,赖伟.日本沼虾生理生态学研究1.温度和体重对其代谢的影响[J].海洋与湖沼,1994,25(3):233-237.
[11]林小涛,颉晓勇,钟金香,等.罗氏沼虾亲虾的能量代谢[J].中山大学学报:自然科学版,2000,39:248-254.
[12]张硕,董双林,王芳.中国对虾生物能量学研究I-温度、体重、盐度和摄食状态对耗氧率和排氨率的影响[J].青岛海洋大学学报,1998,28(2):223-227.
[13]吴立新,刘瑜,王欣,等.饥饿和再投喂对日本囊对虾代谢率的影响[J].大连水产学院学报,2007,22(2):109-112.
[14]雷衍之,杨凤.化学实验[M].北京:中国农业出版社,2004: 152-194.
[15]周洪琪.中国对虾亲虾的能量代谢研究[J].水产学报,1990,14(2):114-119.
[16]董双林,堵南山,赖伟.日本沼虾生理生态学研究Π.温度和体重对能量收支的影响[J].海洋与湖沼,1994,25(3):238-242.
[17]CAWTHORNE D F,BEARD T,DAVENPORT J.Response of juvenilePenaeusmonodonFabricius to natural and artificial sea waters of lowsalinity[J].Aquaculture,1983,32:165-174.
[18]LINDA H M ,LINDA L F.Osmotic and Ionic Regulation.The Biology of Crustacean(5)[M].New York,London:Academic Press,1983:53-161.
[19]维诺格拉多夫M E.海洋生物结构(第1卷)[M].李世珍,杨德渐,译.北京:海洋出版社,1985:23-34.
[20]DALLA V.Salinity response in brick is water populations of the freshwater shrimpPalaemonetesantennarius[J].Oxygen Consumption Comp Biochem Physiol,1987,87A(2):471-478.
[21]施正峰,梅志平,罗其智.日本沼虾能量收支和利用效率的初步研究[J].水产学报,1994,18(3): 191-197.
[22]董双林,堵南山,赖伟.日本沼虾生理生态学研究-PH,Ca2+和NaCl对耗氧率和氨排泄的影响[C]//中国动物学会成立60周年论文集.北京:中国科学技术出版社,1994:176-182.
[23] SPAARGAREN D H,RICHARD P,CECCALDI H J.Excretion of nitrogenous products byPenaeusjaponnicusBate in relation to enviromentalo smotic conditons[J].Comp Biochem Physiol,1982,72A:673-678.
[24] CHEN Jianchu,NAU Fanhua.Oxygen consumption and ammonia Nexcretion ofPenaeuschinensisjuveniles at different salinity levels(Decapod,Penaeidae)[J].J Crusta Biol,1995,15(3): 434-443.
[25] LEI C H,HISEH L Y,CHEN C K.Effects of salinity on the oxygen consump tion and ammnia Nexcretion of young juvenijes of the grass shrimpPenaeusmonodonFabricius[J].Bull Inst Zool Academic Sinica,1989,28: 245-256.
[26]NELSON S G,KNIGHT A W,LI H W.The metabolic cost of food utilization and ammonia producton by juvenileMacrobrachiumrosenberg(Crucea: Palaemonidae)[J].Comp Biochem Physiol,1977,57A: 67-72.
猜你喜欢
世界建筑(2022年11期)2022-12-05 06:56:02
当代水产(2022年8期)2022-09-20 06:46:22
知识窗(2022年6期)2022-07-08 23:40:36
工会博览(2022年9期)2022-06-30 09:30:48
当代水产(2022年5期)2022-06-05 07:55:46
中老年保健(2021年10期)2021-11-30 09:34:06
当代水产(2021年8期)2021-11-04 08:49:04
当代水产(2021年4期)2021-07-20 08:10:54
当代水产(2021年1期)2021-03-19 05:17:14
当代水产(2019年9期)2019-10-08 08:02:44
