
固态发酵菜籽粕对肉仔鸡生长性能、免疫功能及消化酶活性的影响
2012-03-28 10:35胡永娜王之盛李爱科
动物营养学报 2012年7期
胡永娜 王之盛* 李爱科
菜籽粕富含蛋白质和氨基酸,是很好的植物性饲料蛋白质资源[1]。但菜籽粕中由于硫代葡萄糖甙、单宁、植酸等抗营养因子的存在,影响了饲料的安全性、适口性和营养价值利用率,从而限制了其在畜禽饲粮中的使用[2-4]。菜籽粕经微生物固态发酵后,可降低粗纤维含量,有效去除抗营养因子,改善其适口性,同时产生小分子量的肽类物质和益生菌,调节畜禽机体胃肠道微生态平衡,并提高消化酶活性。另外,微生物蛋白质本身也是一种优质的蛋白质资源,可以提高饲料中蛋白质的含量,使其氨基酸组成更加合理。然而,发酵菜籽粕的饲用价值并没有充分体现出来。目前,关于添加发酵菜籽粕对动物生长性能影响的研究已有少量报道。吴文明等[5]在断奶仔猪上的研究发现,发酵菜籽粕替代部分豆粕能提高仔猪平均日增重,降低料重比,降低仔猪腹泻率。蒋玉琴等[6]将发酵菜籽粕按不同比例添加到粉碎的鸡饲料中,研究其对三黄鸡生长性能的影响,结果表明,添加比例在15%时,鸡的平均日增重与对照组(不添加脱毒饼粕)基本接近,余勃等[7]和 Chiang等[8]也有过类似的研究报道。而发酵菜籽粕对肉仔鸡的免疫功能和消化酶活性方面的研究鲜有报道。因此,本研究拟采用混菌发酵处理菜籽粕,通过对肉仔鸡生长性能、免疫功能及消化酶活性相关指标的分析,以探讨微生物处理菜籽粕的应用价值。
1 材料与方法
1.1 试验设计
选用1日龄健康爱拔益加(AA)肉仔鸡180羽,按照初始体重无差异原则将其随机分为3个组,每组6个重复,每个重复10羽。饲养试验时间为42 d,分为试验前期(1~21日龄)和试验后期(22~42日龄)。豆粕对照组饲喂基础饲粮,菜籽粕组用未发酵菜籽粕等氮替代基础饲粮中25%的豆粕,发酵菜籽粕组用固态发酵菜籽粕等氮替代基础饲粮中25%的豆粕。
1.2 试验动物及材料
AA肉仔鸡由北京华都肉鸡场提供。固态发酵菜籽粕在实验室前期试验筛选的基础上,采用混菌发酵,试验所用发酵菌株为本实验室保存菌株CG1.1001、CG1.3001和CG4.1001,分别为枯草芽孢杆菌(Bacill us subtilis)、粪肠球菌(Enter ococcus f aecalis)和产朊假丝酵母(Candida utilis),3株菌株的比例为1∶1∶2。原菜籽粕产自湖北武汉,其底物组成为80%菜籽粕+10%麸皮+10%玉米粉,水料比为1∶1,总接种量为20%。混合物被装入发酵罐后在(30±2)℃静止发酵3 d。发酵前后菜籽粕营养成分和抗营养因子含量的变化见表1。

表1 发酵前后菜籽粕营养成分(干物质基础)Table 2 Nutrient co mposition in r apeseed meal and f er mented r apeseed meal(DM basis) %
1.3 试验饲粮
饲粮配制参考美国NRC(1994)肉鸡营养需要和中国肉鸡饲养标准(2004),按1~21日龄和22~42日龄2个阶段配制饲粮,基础饲粮为玉米-豆粕型饲粮。试验饲粮组成及营养水平见表2。
1.4 饲养管理
试验在国家粮食局科学研究院中试基地进行。所有试验鸡采用立体重叠式3层笼养,分阶段饲养。全程采用粉料,从进场就开始进入试验。进鸡雏前1周,将鸡舍及设备进行严格清洗和消毒。采用空调、红外灯和风扇控温。第1周室温保持在33℃左右,每周降2℃,21日龄后鸡舍温维持在20℃,相对湿度控制在65%~67%。采用连续光照,自然通风,整个试验期间自由采食和饮水。定期打扫圈舍卫生。参照常规免疫程序进行免疫接种。
1.5 测定指标与方法
1.5.1 生长性能
分别于饲养肉仔鸡的1、21和42日龄,以重复为单位称鸡只空腹重,同时记录每个重复投料量和剩料重,计算2阶段的平均日增重(ADG)、平均日采食量(ADFI)和料重比(F/G)。记录腹泻与死亡情况,并在21和42日龄末计算各组的腹泻率与死淘率。
1.5.2 免疫器官指数
分别于21和42日龄从每组中随机抽取6只鸡屠宰后,摘取脾脏、胸腺和法氏囊,称鲜重以计算重量指数。

1.5.3 消化酶活性
21和42日龄时,采集屠宰鸡只,收集十二指肠、空肠、回肠和盲肠的内容物,-20℃保存。在未解冻前用分析天平称重,然后按质量体积比为1∶1的比例加入4℃的生理盐水,在低温(冰浴中)下匀浆30 s,然后用低温离心机将匀浆液在2 500 r/min、4℃条件下离心10 min,取上清液若干后等分,-20℃条件下保存待测。蛋白酶的活性测定采用行业标准SB/T 10317—1999的方法,淀粉酶和脂肪酶的活性用试剂盒测定,试剂盒购自南京建成生物工程研究所。

表2 试验饲粮组成及营养水平(风干基础)Table 2 Co mposition and nutrient levels of experi mental diets(air-dry basis) %
1.6 数据处理
试验数据经过Excel 2003处理后,利用SPSS 13.0统计软件中的one-way ANOVA程序进行单因素方差分析,差异显著时采用Duncan氏法进行多重比较,试验结果以平均值±标准差表示。
2 结 果
2.1 发酵菜籽粕对肉仔鸡生长性能的影响
由表3可知,无论是在饲养前期(1~21日龄)还是后期(22~42日龄),发酵菜籽粕组肉仔鸡的平均日采食量和平均日增重均优于豆粕对照组和菜籽粕组(P>0.05),发酵菜籽粕组的料重比低于菜籽粕组,与豆粕对照组差异不显著(P>0.05),但各组肉仔鸡的平均日增重、平均日采食量和料重比均无显著差异(P>0.05)。发酵菜籽粕组较菜籽粕组可显著提高肉仔鸡全期(1~42日龄)的平均日增重3.73%(P<0.05)。

表3 发酵菜籽粕对肉仔鸡平均日增重、平均日采食量及料重比的影响Table 3 Eff ects of fer mented rapeseed meal on ADG,ADFI and F/G of br oilers
由表4可知,在试验前期和试验后期,发酵菜籽粕组的死淘率低于其他组,但差异不显著(P>0.05);从腹泻率来看,发酵菜籽粕组低于豆粕对照组和菜籽粕组,差异不显著(P>0.05)。
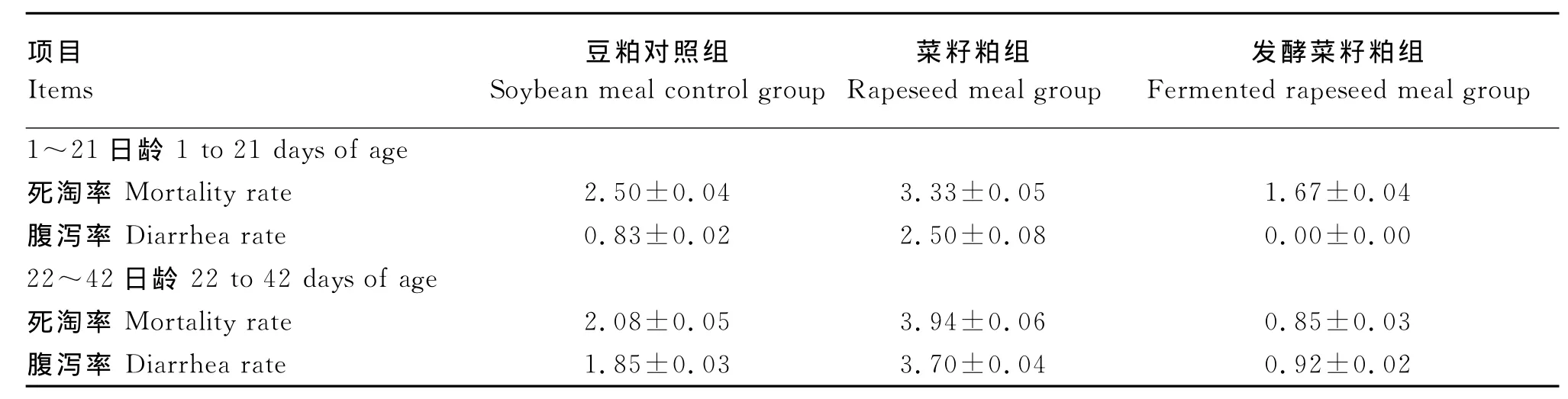
表4 发酵菜籽粕肉仔鸡死淘率及腹泻率的影响Table 4 Effects of fer mented rapeseed meal on mortality and diarrhea rate of broilers %
2.2 发酵菜籽粕对肉仔鸡免疫器官的影响
由表5可知,21日龄发酵菜籽粕组提高了胸腺、脾脏和法氏囊指数。发酵菜籽粕组的肉仔鸡胸腺和脾脏指数分别显著高于菜籽粕组28.28%和36.58%(P<0.05),发酵菜籽粕组的法氏囊指数高于豆粕对照组和菜籽粕组,但差异不显著(P>0.05);42日龄菜籽粕组的肉仔鸡脾脏指数显著高于豆粕对照组51.00%(P<0.05),而3组的胸腺和法氏囊指数差异不显著(P>0.05)。
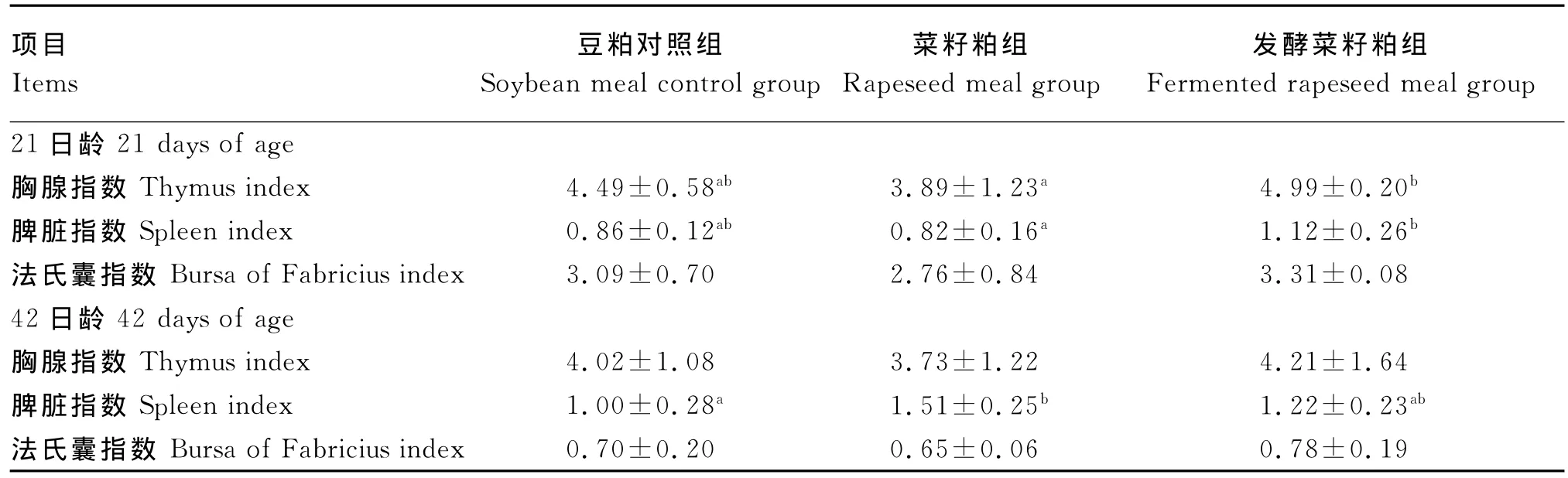
表5 发酵菜籽粕对肉仔鸡免疫器官的影响Table 5 Eff ects of fer mented rapeseed meal on immune organs of broilers g/kg
2.3 发酵菜籽粕对肉仔鸡消化酶活性的影响
由表6可知,21日龄时,与豆粕对照组相比,菜籽粕组十二指肠蛋白酶、淀粉酶和脂肪酶活性分别显著降低了5.58%、2.36%和8.40%(P<0.05),发酵菜籽粕组十二指肠蛋白酶、淀粉酶和脂肪酶活性分别显著提高了5.00%、3.04%和15.11%(P<0.05);菜籽粕组空肠蛋白酶、淀粉酶和脂肪酶活性较豆粕对照组分别显著降低了6.81%、2.64%和5.35%(P<0.05),发酵菜籽粕组较豆粕对照组空肠蛋白酶、淀粉酶和脂肪酶活性分别显著提高了4.12%、3.56%和18.30%(P<0.05);与豆粕对照组相比,菜籽粕组回肠蛋白酶、淀粉酶和脂肪酶活性分别显著降低了6.53%、2.59%和7.69%(P<0.05),发酵菜籽粕组空肠蛋白酶、淀粉酶和脂肪酶活性分别显著提高了4.62%、4.28%和15.03%(P<0.05);与豆粕对照组相比,菜籽粕组盲肠蛋白酶、淀粉酶和脂肪酶活性分别显著降低了7.17%、2.78%和4.96%(P<0.05),发酵菜籽粕组盲肠蛋白酶、淀粉酶和脂肪酶活性分别显著提高了4.23%、4.29%和17.58%(P<0.05)。
42日龄时,菜籽粕组十二指肠蛋白酶、淀粉酶和脂肪酶活性较豆粕对照组分别降低了6.67%(P<0.05)、1.32%(P<0.05)和5.77%(P>0.05),发酵菜籽粕组较豆粕对照组十二指肠蛋白酶、淀粉酶和脂肪酶活性分别显著提高了5.96%、4.33%和12.98%(P<0.05);菜籽粕组空肠蛋白酶、淀粉酶和脂肪酶活性较豆粕对照组分别显著降低了5.48%、2.55%和6.46%(P<0.05),发酵菜籽粕组空肠蛋白酶、淀粉酶和脂肪酶活性较豆粕对照组分别显著提高了3.91%、3.73%和21.09%(P<0.05);与豆粕对照组相比,菜籽粕组回肠蛋白酶、淀粉酶和脂肪酶活性分别显著降低了6.66%、2.32%和0.71%(P<0.05),发酵菜籽粕组回肠蛋白酶、淀粉酶和脂肪酶活性分别显著提高了4.15%、4.73%和15.49%(P<0.05);与豆粕对照组相比,菜籽粕组盲肠蛋白酶、淀粉酶和脂肪酶活性分别显著降低了6.75%、1.70%和4.35%(P<0.05),发酵菜籽粕组盲肠蛋白酶、淀粉酶和脂肪酶活性分别显著提高了3.56%、5.67%和19.26%(P<0.05)。
3 讨 论
3.1 发酵菜籽粕对肉仔鸡生长性能的影响
许多研究报道表明,影响菜籽粕在家禽饲料中使用的限制因素主要是对畜禽有毒副作用的硫代葡萄糖甙、单宁和植酸等抗营养物质,从而在很大程度上制约了其在饲料中的应用[9]。本研究利用混菌固态发酵菜籽粕有效地降低硫代葡萄糖甙等抗营养因子的同时,提高了菜籽粕的粗蛋白质和小肽含量,改善了菜籽粕的营养价值和适口性,这与 Vig等[10]和孙宏[11]的报道一致。研究显示,菜籽粕经过发酵处理,使其中的抗营养因子去除或减少,能有效提高动物的生长性能[5]。本研究结果表明,饲粮中固态发酵菜籽粕对肉仔鸡1~21日龄、22~42日龄及1~42日龄的平均日采食量和料重比均无显著影响,这与余勃等[7]的结论相一致,发酵菜籽粕还降低了肉仔鸡的腹泻率和死淘率,但差异不显著。而未发酵菜籽粕组降低了采食量,增加了死淘率,影响了肉仔鸡的生长性能,这与 Mc Neill等[12]的研究相一致。发酵菜籽粕组肉仔鸡全期的平均日增重显著高于未发酵菜籽粕组,这表明除采食量外,代谢能、蛋白质和氨基酸的生物利用率及硫代葡萄糖甙含量也是引起肉仔鸡生长性能的关键,而发酵菜籽粕降解产生的小肽易于消化吸收,可能对肉仔鸡生长性能具有改善作用。
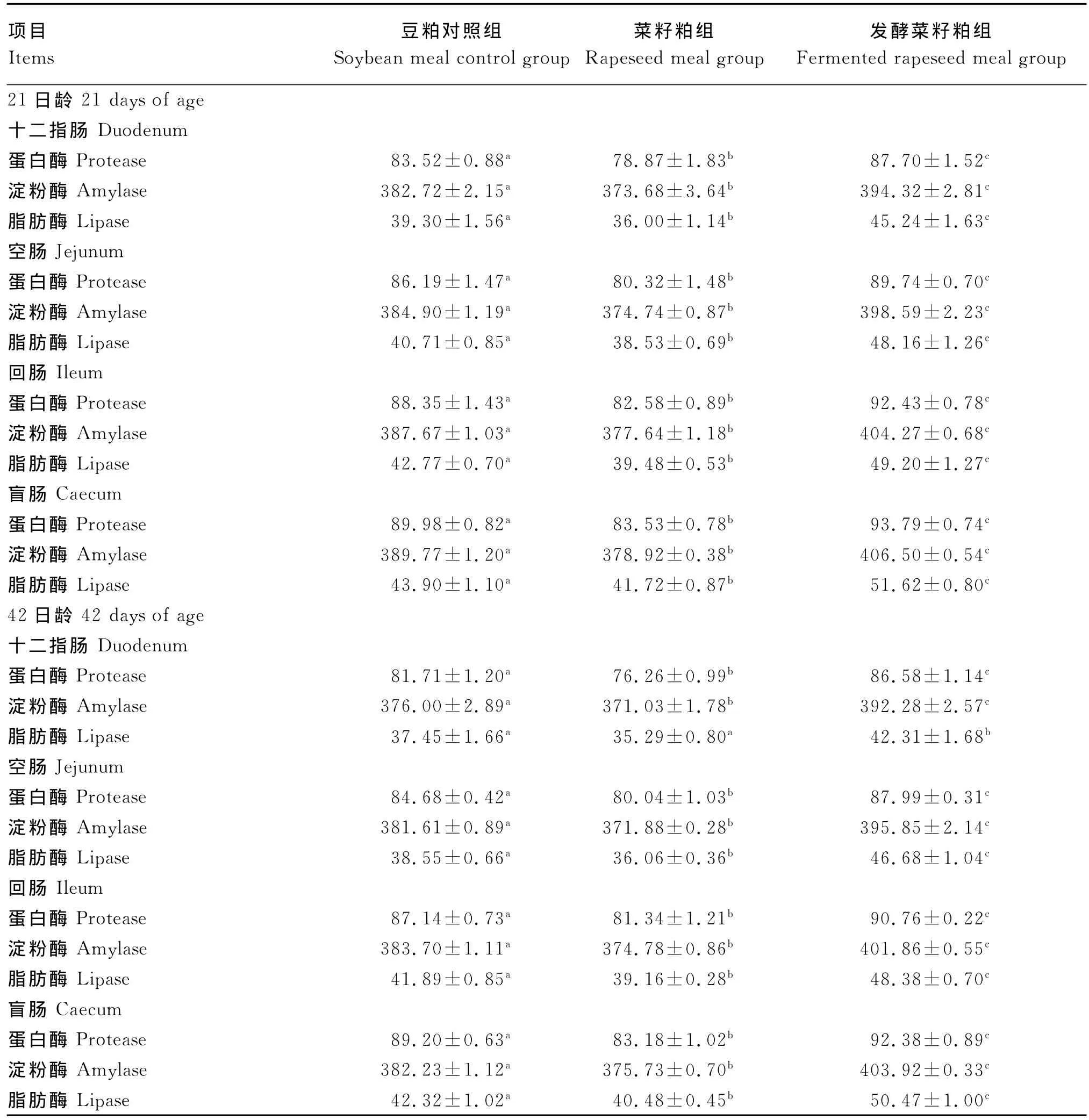
表6 发酵菜籽粕对肉仔鸡消化酶活的影响Table 6 Effects of fer mented rapeseed meal on digestive enzy me activity of broilers U/g
3.2 发酵菜籽粕对肉仔鸡免疫功能的影响
肉仔鸡脾脏、胸腺和法氏囊指数反映机体主要免疫器官的生长发育状况,是从免疫器官发育的角度评价机体免疫状态的指标。免疫器官的发育状况直接影响机体免疫应答水平和抵抗外来微生物感染和入侵的能力,其绝对重量和相对重量的增加或降低,可表明机体的细胞免疫和体液免疫机能增强或降低[13-15]。由本试验结果来看,无论是在前期还是后期发酵菜籽粕对法氏囊指数均无显著影响,发酵菜籽粕能够显著提高肉仔鸡前期的胸腺指数和脾脏指数,但在肉仔鸡生长后期,胸腺指数和脾脏指数均没有产生显著变化。胸腺是家禽的一级淋巴器官,而脾脏则是家禽体内最大的淋巴器官,因此肉仔鸡的胸腺和脾脏相对重量增加,意味着发酵菜籽粕可以刺激肉仔鸡的免疫应答反应,促进肉仔鸡免疫器官的发育,增强肉仔鸡的免疫功能,这在发酵菜籽粕应用试验中尚未见报道。本试验制备的发酵菜籽粕的活菌数达到1.4×109CFU/g,Li等[16]曾证实,芽孢杆菌能够刺激肉仔鸡胸腺指数、脾脏指数和法氏囊指数的增加,因此推断胸腺指数和脾脏指数的增加可能与菜籽粕发酵后产生了芽孢杆菌等有益微生物有关,对此有待于进一步研究。
3.3 发酵菜籽粕对肉仔鸡消化酶活性的影响
消化酶是动物消化利用营养物质的首要工具,其活性高低直接影响到动物对饲料的利用程度[17]。十二指肠、空肠、回肠和盲肠内容物中的蛋白酶、淀粉酶和脂肪酶活性的变化是反映机体对蛋白质、淀粉和脂肪消化能力的一个重要指标[18]。本试验研究发现,发酵菜籽粕可显著提高肉仔鸡十二指肠、空肠、回肠和盲肠的蛋白酶、淀粉酶和脂肪酶活性。发酵菜籽粕能提高肉仔鸡肠道消化酶活性,可能是以下几方面的原因:1)菜籽粕经发酵处理后,其中的硫代葡萄糖甙等抗营养因子已经大部分被去除,肉仔鸡采食进入消化道后,对肠道不会造成过敏反应和损伤,从而保证消化酶的正常活性[19];2)增加了肉仔鸡小肠各段绒毛高度,降低了隐窝深度,有效地维持了肠绒毛的光滑平整状态,提高了消化酶活性;3)发酵产生的小肽类物质能显著增加肠道食糜中蛋白酶、脂肪酶和淀粉酶等消化酶的分泌与活性[20]。发酵菜籽粕中乳杆菌分泌的乳酸能激活酸性蛋白酶活性,在消化过程中与体内的酶起到协同作用,也可使肠道环境偏酸性,有利于刺激肠壁消化腺的代谢分泌活动,增加消化酶的分泌量,提高消化酶的活力。但具体的作用机制还有待进一步研究。
4 结 论
①固态发酵菜籽粕等氮替代基础饲粮中25%的豆粕对肉仔鸡全期的平均日采食量无显著影响。
②固态发酵菜籽粕等氮替代基础饲粮中25%的豆粕可明显提高肉仔鸡的免疫能力和肠道消化酶活性,有利于提高肉仔鸡全期平均日增重,促进肉仔鸡的生长。
[1] 席鹏彬,马永喜.中国菜籽饼粕化学组成特点及其影响因素的研究[J].中国畜牧杂志,2004,40(10):12-15.
[2] CAMPBELL L D,SMITH T K.Response of growing chicks to high dietar y content of rapeseed meal[J].Britain Poultry Science,1979,20:231-237.
[3] ELANGOVAN A V,VERMA V S,SASTRYV R B,et al.Eff ect of feeding high glucosinolate rapeseed meal to laying Japanese quail[J].Asian-Australasian Jour nal Ani mal Science,2001,14:1304-1307.
[4] SUN Z D,ISMOND M A H.Study on the extraction,isolation of canola meal protein after removing the phenolic co mpounds[J].Jour nal of Huazhong Agricultural University,1999,18(4):394-396.
[5] 吴文明,张丽娟.发酵菜粕对仔猪生产性能影响的试验研究[J].饲料广角,2010,2:23-24.
[6] 蒋玉琴,李荣林.复合菌脱毒菜籽饼粕及其应用Ⅱ:日粮添加脱毒菜粕对三黄鸡生长的影响[J].江苏农业学报,2001,17(1):52-55.
[7] 余勃,游金明.固态发酵菜粕替代日粮中豆粕对肉仔鸡生长性能的影响[J].动物营养学报,2009,21(2):239-244.
[8] CHIANG G,LU W Q,PIAO X S,et al.Effects of feeding solid-state f er mented rapeseed meal on perf or mance,nutrient digestibility,intestinal ecology and intestinal morphology of broiler chickens[J].Asian-Australasian Jour nal Ani mal Science,2010,23(2):263-271.
[9] TRIPATHI M K,MISHRA A S.Glucosinolates in animal nutrition:a review[J].Ani mal Feed Science and Technology,2006,132:1-27.
[10] VIG A P,WALIA A.Beneficial effects of Rhizopusoligospor us f er mentation on reduction of glucosinolates,fibre and phytic acid in r apeseed (Brassica napus)meal[J].Bioresource Technology,2001,78(3):309-312.
[11] 孙宏.微生物发酵法对菜粕脱毒及蛋白质改良的研究[D].硕士学位论文.武汉:华中农业大学,2009:60-64.
[12] MCNEILL L,BEMARD L K,MACLEOD M G.Food intake,gr owth rate,f ood conversion and f ood choice in br oilers fed on diets high in r apeseed meal and pea meal with observations of the resulting poultry meat[J].Britain Poultry Science,2004,45:519-523.
[13] STUKIE P D,BENZO C A.Avian physiology[M].New Yor k:Springer-Verlag,1986.
[14] KATANBAF M N,DUNNINGTON E A,et al.Re-stricted f eeding in early and late-f eathering chickens[J].Poultry Science,1989,68:344-351.
[15] 何涛.棉籽粕的发酵脱毒及其在肉仔鸡中的应用研究[D].硕士学位论文.北京:中国农业科学院,2008:44.
[16] LI S P,ZHAO X J,WANG J Y.Synergy of astragalus polysaccharides and pr obiotics(Lactobacillus and Bacillus cereus)on i mmunity and intestinal micr obiota in chicks[J].Poultry Science,2009,88(2):519-525.
[17] 罗辉,剑叶华.微生态制剂对鱼类肠道结构和消化酶活性的影响[J].水产科学,2006,25(2):105-108.
[18] 杨凤.动物营养学[M].北京:中国农业出版社,1999:18-21.
[19] 刘海燕,闫晓刚.发酵酶解豆粕对小鼠血液抗氧化指标和胃肠道消化酶活性及肠道菌群的影响[J].中国畜牧杂志,2011,47(21):41-44.
[20] 王恬,傅永明,吕俊龙,等.小肽营养素对断奶仔猪生产性能及小肠发育的影响[J].畜牧与兽医,2003,35(6):4-8.
猜你喜欢
CHINA TODAY(2022年4期)2022-11-22
当代水产(2021年9期)2021-12-02
养殖与饲料(2021年5期)2021-11-30
现代畜牧科技(2021年10期)2021-11-19
当代水产(2021年8期)2021-11-04
花卉(2021年17期)2021-09-15
医学前沿(2021年18期)2021-04-14
兽医导刊(2019年20期)2019-08-19
今日畜牧兽医(2018年9期)2018-02-13
北方牧业(2016年6期)2016-12-17
