
整联蛋白及其信号转导途径对骨骼肌生长发育的调控
2012-03-14 05:33李欣蔚
动物营养学报 2012年7期
李欣蔚 张 勇
(沈阳农业大学畜牧兽医学院,沈阳110866)
整联蛋白(integrin)是由α和β2个非共价键结合的跨膜亚基组成的异源二聚体,α和β亚基均由氨基酸序列较长的胞外区、跨膜区和氨基酸序列较短的胞质区组成(只有β4亚基有长的胞质区),α和β链的氨基末端形成的球形区域部分为胞外配体结合域[1]。有研究表明,在果蝇中,整联蛋白在细胞黏附、细胞迁移、细胞增殖和细胞凋亡等过程中同样发挥重要作用[2-3]。同时,Elizabeth等[4]指出,整联蛋白黏着复合物会促进特定肌肉组织的发育,主要是通过促进蛋白质合成以及加强细胞原有功能实现的。整联蛋白通过介导细胞外基质,构成整联蛋白信号通路来完成对细胞凋亡、细胞增殖以及细胞迁移的信号转导调控[5]。
1 整联蛋白配体的生物学特性及其信号通路的特点
1.1 整联蛋白配体的结构和生物学特性
由于α亚基具有18种不同构型,而β亚基具有8种不同构型,故整联蛋白可以组成24种α、β异源二聚体复合物。多数α亚基只能与1种β亚基结合构成异源二聚体,但有些α亚基可与几种不同的β亚基组合;而大部分β亚基则可以结合多种不同的α亚基;同时,β1、β3、β4、α3、α6、αⅡb等亚基的基因mRNA分子可有不同的剪接形式。以上诸多因素都增加了整联蛋白分子的多样性。
α和β亚基均由胞外区和胞浆区组成,胞外区和胞浆区共含有700~1 000个氨基酸。α亚基的胞外区由Ⅰ结构域、β-螺旋桨(β-propeller)、thigh、Calf-1、Calf-2构成。而β亚基(β1~β8)包含8个胞外区,即Ⅰ结构域、hybrid、神经丛蛋白-脑信号蛋白-整联蛋白(plexin-semaphorin-integrin,PSI)子域、β-尾区和4个类表皮生长因子(EGF-like)子域[6]。以整联蛋白β1为例,如图1[7]所示:整联蛋白β1共含有831个氨基酸,其信号肽含有20个氨基酸残基,位于氨基酸1~20;胞外区域的氨基端部分(INB)是保守子域,共含有449个氨基酸残基,位于氨基酸37~485;i-like子域位于INB区域内,是A型超家族子域中的成员,该家族含有一种原型分子,叫做von willebrand因子;金属离子依赖性黏附位点(metal ion dependent adhesion site,MIDAS)氨基酸序列在173~177;PSI子域位于氨基酸31~73;EGF-like子域位于氨基酸513~524和607~618。其中,MIDAS存在于β亚基和部分α亚基的Ⅰ结构域,并位于具有调节作用的Ca2+结合位点附近,Mg2+、Mn2+和Ca2+均可以与 MIDAS相互作用使得整联蛋白构象改变,从而导致其与配体的结合或解离[8]。
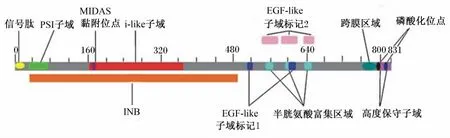
图1 整联蛋白β1的氨基酸序列及保守子域Fig.1 Amino acid sequence and conserved domains of integrinβ1[7]
1.2 整联蛋白及其信号转导途径的活化特点
整联蛋白作为介导信号传递的膜分子,是通过独特的途径转导信号来调节细胞的各种生物学功能的。而整联蛋白的活化主要通过触发由细胞内到细胞外的信号转导,从而实现与细胞外配体的低亲和力状态到高亲和力状态的改变,即整联蛋白信号系统。而最新研究发现,动物受到应激后,激活的细胞外信号调节激酶(extracellular regulated kinase,ERK)会移动到细胞核表面激活整联蛋白系统[9]。
在整联蛋白的活化过程中,小鼠黏着斑蛋白-3(kindlin-3)基因敲除的试验证实:kindlin-3才是整联蛋白活化所必需的[10-12]。同时,黏着斑蛋白-1(kindlin-1)可与整联蛋白β1和整联蛋白β3胞质区作用,而kindlin-3能与白细胞特异性的整联蛋白β2胞质区结合[11]。有研究表明,尽管多种蛋白质都可以与β整联蛋白胞质区发生相互作用,但此前只有踝蛋白被确切证实能够触发整联蛋白的活化[13],而黏着斑蛋白(kindlin)的F3子域中的TxxNxxY序列是其主要的结合位点[14]。当整联蛋白的β亚基的胞质尾区与F3子域结合后,破坏了α、β之间的盐桥,继而触发α、β跨膜构象转变为高亲和力构象[15]。
整联蛋白系统被活化后,黏着斑激酶(focal adhesion kinase,FAK)是胞内第1个信号分子,首先进行磷酸化。FAK广泛分布于血小板、内皮细胞、中性粒细胞和单核细胞等细胞的胞质中,在细胞迁移中发挥作用[16-19]。继而由整联蛋白连接激酶(integrin-linked kinase,ILK)作为蛋白质间相互作用网络中的集结点,通过其偶联的信号通路,在调节细胞生物学功能上发挥多种效应。具体信号通路是:当整联蛋白依赖的细胞黏附激活磷酸肌醇-3激酶(PI3K),其产物多磷酸肌醇(PIP)可激活ILK[20]。
2 由整联蛋白系统介导的ERK信号通路的生物学功能
整联蛋白不仅可以通过识别细胞外基质的精氨酸-甘氨酸-天冬氨酸(Arg-Gly-Asp,RGD)序列介导细胞与细胞外基质之间的黏附,与免疫球蛋白超家族分子结构介导细胞与细胞间的黏附,还可以双向转导细胞内外的信号,直接影响细胞的生存、生长、增殖、分化以及凋亡等生物学行为。Fong等[21]研究发现,骨桥蛋白可以识别整联蛋白αⅤβ3,启动FAK/激活磷酸肌醇3(PI3)/ERK细胞内信号通路,进而促进细胞迁移。Roman等[22]以及Shibue等[23]也指出,整联蛋白β1可以通过细胞外信号调节激酶1(ERK1)/细胞外信号调节激酶2(ERK2)信号通路上游的FAK调控细胞的增殖,通过抑制活化的整联蛋白α3,利用FAK/ERK/核转录因子κB(NFκB)信号途径调控基质金属蛋 白 酶 (matrix metalloproteinase,MMP)的表达。
此外,在硫酸乙酰肝素蛋白聚糖(heparan sulfate proteoglycans,HSPGs)介导细胞分布以及维持细胞骨架硬度的过程中,整联蛋白通过下调FAK/ERK信号通路来发挥重要的动力转导作用。如图2[24]所示,HSPGs可以通过与整联蛋白调控的细胞基质黏附复合物共区域化,或者通过皮层肌动蛋白细胞骨架上的黏附复合物相互作用,或者同时发 挥 以 上 2 种 作 用 来 激 活 FAK/ERK/c-Jun/MMP-13信号中枢。活化后的信号通路不仅可以调控MMP-13的表达,而且有研究指出,在皮肤损伤修复以及成肌纤维细胞的分化过程中,通过胞间质的作用,整联蛋白也发挥了重要的动力转导作用[25]。
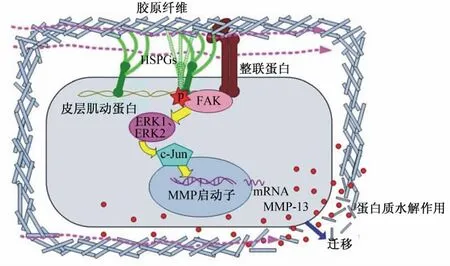
图2 整联蛋白在FAK/ERK信号通路中的动力转导作用Fig.2 The mechanotransduction of integrin in FAK/ERK signal pathway[24]
3 整联蛋白及其信号转导途径对动物肌肉生长发育的调控
3.1 整联蛋白对动物肌肉生长发育的调控
骨骼肌的生长发育是一个十分复杂的过程,涉及大量基因的表达及网络式调控[26]。而骨骼肌生长速度最终取决于骨骼肌细胞的数量、肌肉蛋白质合成和降解速度[27]。在骨骼肌发育过程中,肌肉组织之所以能够构建成为可伸展的、可受力的分生组织,是因为整联蛋白及其调控的细胞黏着力提供了结构上的稳定性。同样的,有研究表明,在骨骼肌形成过程中,整联蛋白集中在Z线,整联蛋白缺乏将会导致Z线发育缺陷[28]。Perkins等[29]的试验表明,骨骼肌中的整联蛋白调控的黏着力下降会导致肌肉功能的逐渐缺失,原因在于肌节细胞结构无法维持。而这种肌肉功能上的缺失是渐进的,随着年龄的增长,肌节肌动蛋白、Z线和M线会出现解离。电子显微镜显微分析显示,由于整联蛋白在黏着力调控过程中发生的还原反应,细丝蛋白会与Z线解离,随后Z线与细胞膜逐渐分离,最终导致Z线的分解。与此同时,整联蛋白可作为骨骼肌层粘连蛋白网络调控的细胞膜受体,参与骨骼肌细胞骨架的形成和细胞内信号途径的介导[30]。
3.2 由泛素介导的整联蛋白下游存活信号通路对肌肉发育的影响
泛素(ubiquitin)是一类高度保守的蛋白质,含有76个氨基酸,分子质量大约为8 500u。泛素家族除了保持有限序列的同源性以外,所有的泛素家族成员都具有典型的β-grasp泛素折叠,这也显示出泛素家族结构上的同源性[31]。泛素存在于大多数真核细胞中,它的主要功能是标记需要分解掉的蛋白质,使其被水解。当附有泛素的蛋白质移动到桶状的蛋白酶的时候,蛋白酶就会将蛋白质水解。有研究表明,整联蛋白β1的活化作用和泛素的表达均可以激活PI3K/蛋白激酶B(Akt)通路来进一步抑制细胞凋亡[32]。
在肌间盘和肋节间,整联蛋白β1与多种蛋白质如骨骼肌特异蛋白质(muscle-specific protein,melusin)相衔接(图3[33])。β肾上腺素受体刺激成人的心肌细胞,继而引起糖原合成酶激酶3β(glycogen synthase kinase 3β,GSK3β)和c-Jun氨基端激酶(JNK)的活化,在整联蛋白β1和基质金属蛋白酶-2(matrix metalloproteinase-2,MMP-2)的参与完成下游信号的传导,引起细胞凋亡。另有研究表明[34],阻止 MMP-2的活化会导致肌细胞骨架重组受抑制,同时引起肌细胞功能性异常。同样的,切断melusin基因后也会抑制Akt介导的GSK3β的磷酸化作用,继而抑制了细胞凋亡。同时,泛素介导的信号通路中所涉及的诸多蛋白质分子,将会被肌细胞骨架所重新吸收,然后完成下游信号通路的转导。在β肾上腺素受体的刺激下,整联蛋白β3不会被心肌细胞的闰盘泛素化,而在这段时间,酪氨酸激酶活化后的下游因子——黏着斑复合体(focal adhesion complex,FAC)逐渐形成。所以,在β肾上腺素受体的刺激下,整联蛋白β1和整联蛋白β3都会引起肌细胞的代偿性生长,整联蛋白与其他蛋白质共同形成了网络式的级联反应,而在这些级联反应中,由泛素介导的整联蛋白β1下游存活信号通路会被不断激活。
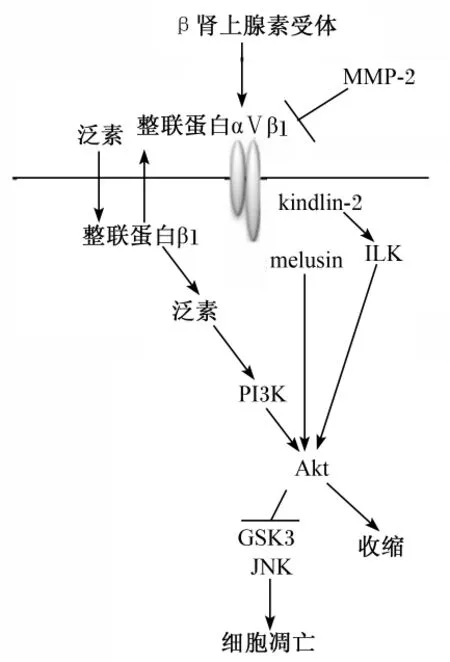
图3 由泛素介导的整联蛋白β1下游存活通路Fig.3 Ubiquitin-mediated survival signaling downstream of integrinβ1[33]
4 小 结
整联蛋白是一类细胞表面受体分子家族,广泛存在于各种组织中,可使细胞黏附于胞外基质并介导来自基质的机械信号和化学信号,具有激活胞浆激酶和维持生长因子的生物活性,实现细胞外基质-整联蛋白-细胞内的信号转导功能。而骨骼肌生长发育需要蛋白质合成和降解之间的平衡,涉及合成代谢与分解代谢之间的比例关系。因此,根据整联蛋白信号通路的特点可以推测,在肌肉蛋白质降解和合成代谢过程中,整联蛋白及其信号通路很可能发挥重要作用。目前,对于整联蛋白的研究尚处于人类和动物疾病的预防及治疗阶段,在肌肉发育以及动物肉品质改良方面甚少。所以,深入研究整联蛋白及其信号通路在肌肉发育方面的调控机理,便可以通过调节整联蛋白信号途径来进一步改善动物肉品质,可以说,在动物营养领域的研究中,整联蛋白及其信号通路极具研究前景。
[1] LUO B H,CARMAN C V,SPRINGER T A.Structural basis of integrin regulation and signaling[J].Annual Review Immunology,2007,25(6):619-647.
[2] O'REILLYA M,LEE H H,SIMON M A.Integrins control the positioning and proliferation of follicle stem cells in the Drosophila ovary[J].Journal of Cell Biology,2008,182:801-815.
[3] DINKINS M B,FRATTO V M,LEMOSY E K.Integrinαchain exhibit distinct temporal and spatial localization patterns in epithelial cells of the Drosophila ovary[J].Developmental Dynamics,2008,237:3927-3939.
[4] ETHERIDGE T,OCZYPOK E A,LEHMANN S,et al.Calpains mediate integrin attachment complex maintenance of adoult muscle in Caenorhabditis elegans[J].PLoS Gentics,2012,1(8):1-20.
[5] SEJEONG S,LAURA W,SANG-OH Y.Integrin trafficking and tumor progression[J].International Journal of Cell Biology,2012,2012:1-7.
[6] STEWART P L,NEMEROS G R.Cell integrins:commonly used receptors for diverse viral pathogens[J].Trends Microbiol,2007,15(11):500-507.
[7] HU J,ZHAO H F,LIU J,et al.Integrinβ1subunit from Ostrinia furnacalis hemocytes:molecular characterization,express,and effects on the spreading of plasmatocytes[J].Journal of Insect Physiology,2010,56:1846-1856.
[8] WEGENER K L.Structural basis of integrin activation by talin[J].Cell,2007,128(1):171-182.
[9] BHARADWAJ S,VASABTH G,MASUELLI L,et al.Inhibition of nuclear accumulation of phosphorylated ERK by tropomyosin-1-mediated cytoskeletal reorganization[J].Cancer Molecular,2008,4(5):139-144.
[10] MOSER M,NIESWANDT B,USSAR S,et al.Kindlin-3is essential for integrin activation and platelet aggregation[J].Nature Medicine,2008,14(3):325-330.
[11] MOSER M,BAUER M,SCHMID S,et al.Kindlin-2 is required for myocyte elongation and is essential for myogensis[J].BioMed Central Cell Biology,2008,9:36.
[12] LARJAVA H,WU C.Kindlins:essential regulators of integrin signalling and cell-matrix adhesion[J].European Molecular Biology Organization Reports,2008,9(12):1203-1208.
[13] WEGENER K L,PARTRIDGE A W,HAN J,et al.Structural basis of integrin activation by talin[J].Cell,2007,128(1):171-182.
[14] MA Y Q,QIN J,WU C,et al.Kindlin-2 (Mig-2):a co-activator ofβ3integrins[J].Cell Biology,2008,181(3):439-446.
[15] XIONG J P,ALONSO J L.Crystal structure of the complete integrinαⅤβ3ectodomain plus anα/βtransmembrane fragmanet[J].Journal of Cell Biology,2009,186(4):589-600.
[16] JONES M L,SHAWE-TAYLOR A J,WILLIAMS C M,et al.Characterization of a novel focal adhesion kinase inhibitor in human plateles[J].Biochemical and Biophysical Research Communications,2009,389(1):198-203.
[17] TAVORA B,BATISTA S,REYNOLDS L E,et al.Endothelial FAK is required for tumour angiogenesis[J].European Molecular Biology Organization Molecular Medicine,2010,2(12):516-528.
[18] KASORN A,ALCAIDE P,JIA Y,et al.Focal adhesion kinase regulates pathogen-killing capability and life span of neutrophils via mediating both adhesiondependent and-independent cellular signals[J].Journal of Immunology,2009,183(2):1032-1043.
[19] PARK Y M,FEBBRAIO M,SILVERSTEIN R L.CD36modulates migration of mouse and human macrophages in response to oxidized LDL and may contribute to macrophage trapping in the arterial intima[J].Journal of Clinical Investigation,2009,119(1):136-145.
[20] MCDONALD P C,OLOUMI A,MILLS J,et al.Rictor and integrin-linked kinase interact and regulate Akt phosphorylation and cancer cell survival[J].Cancer Research,2008,68(6):1618-1624.
[21] FONG Y C,LIU S C,HUANG C Y,et al.Osteopontin increases lung cancer cells migration via activation of theαⅤβ3integrin/FAK/Akt and NF-κB-dependent pathway[J].Lung Cancer,2009,64(3):263-270.
[22] ROMAN J,RITZENTHALER J D,ROSER-PAGE S,et al.α5β1-integrin expression is essential for tumor progression in experimental lung cancer[J].American Journal Respiratory Cell and Molecular Biology,2010,43(6):684-691.
[23] SHIBUE T,WEINBERG R A.Integrinβ1-focal adhesion kinase signaling direcrts the proliferation of metastatic cancer cells disseminated in the lungs[J].Proceedings of the Nature Academy of Sciences(USA),2009,106(25):10290-10295.
[24] SHI Z D,WANG H,JOHN M,et al.Heparan sulfate proteoglycans mediate interstitial flow mechanotransduction regulating MMP-13expression and cell motility via FAK-ERK in 3Dcollagen[J].PloS One,2011,6(1):1-10.
[25] HUANG C Y,FU X L,LIU J,et al.The involvement of integrinβ1signaling in the migration and myofibroblastic differentiation of skin fibroblasts on anisotropic collagen-containing nanofibers[J].Biomaterials,2012,33:1791-1800.
[26] 张勇,孙璀.钙调磷酸酶-活化T细胞核因子信号途径在骨骼肌细胞生长和发育中生理作用的研究进展[J].动物营养学报,2011,23(4):536-541.
[27] 张勇,邓科.骨骼肌特异性蛋白酶与蛋白质降解[J].动物营养学报,2011,23(4):542-545.
[28] SPARROW J C,SCHOCK F.The initial steps of myofibril assembly:integrins pave the way[J].Nature Reviews Molecular Cell Biology,2009,10:293-298.
[29] PERKINS A D,STEPHANIE J,SHAMSIAN P A I,et al.Integrin-mediated adhesion maintains sarcomeric integrity[J].Developmental Biology,2010,338:15-27.
[30] GAWLIK K I,DURBEEJ M.Skeletal muscle laminin and MDC1A:pathogenesis and treatment strategies[J].Skeletal Muscle,2011(1):2-13.
[31] VERTEGAAL A O.Uncovering ubiquitin and ubiquitin-like signaling networks[J].American Chemical Society,2011,111(12):7923-7940.
[32] SINGH M,ROGINSKAYA M,DALAL S,et al.Extracellular ubiquitin inhibitsβ-AR-stimulated apoptosis in cardiac myocytes:role of GSK-3βand mitochondrial pathways[J].Cardiovascular Research,2010,86(1):20-28.
[33] HARSTON R K,KUPPUSWAMY D.Integrins are the necessary links to hypertrophic growth in cardiomyocytes[J].Journal of Signal Transduction,2011,10:1-8.
[34] MENON B,SINGH M,ROSS R S,et al.β-adrenergic receptor-stimulated apoptosis in adult cardiacmyocytes involves MMP-2-mediated disruption ofβintegrin signaling and mitochondrial pathway[J].American Journal of Physiology,2006,290(1):254-261.
猜你喜欢
电子学报(2022年2期)2022-04-18
煤炭工程(2021年7期)2021-07-27
世界最新医学信息文摘(2020年68期)2020-12-25
医学综述(2020年11期)2020-02-16
延安大学学报(医学科学版)(2019年1期)2019-03-29
物理学报(2018年10期)2018-06-14
化工学报(2016年10期)2016-10-13
浙江大学学报(农业与生命科学版)(2015年4期)2015-12-15
中国医学科学院学报(2015年5期)2015-03-01
现代检验医学杂志(2015年2期)2015-02-06
