
具有HollingⅡ型功能性反应的捕食者-食饵模型的定性分析
2012-03-12 03:38于丽颖
东北电力大学学报 2012年6期
于丽颖
(北方工业学校数学教研室,辽宁盘锦124021)
捕食者-食饵关系是自然界中普遍存在的物种间相互作用的一种基本关系.揭示具有功能性反应的捕食者-食饵相互作用关系,具有非常重要的理论意义和应用价值,许多学者对此作了大量研究工作。1965年,Holling[1]在试验的基础上,对不同类型的物种,提出了三种不同的功能性反应函数,被广泛应用于生物动力系统研究[2-4]。但在已有的工作中,大多假设食饵种群的增长服从经典的Logistic方程

其中,N是t时刻的种群密度,r,k是正常数。Logistic方程假设种群的相对增长率N'/N是种群密度的线性函数,这一假设受到了学者们的质疑[5-7]。
1963年,F E Smith[8]在实验室里利用扩充培养的方法对Daphnia Magna种群进行了研究,发现此种群的相对增长率不满足线性规律,提出了一种形式更为一般、更符合实际的单种群增长模型(称Smith模型或食物有限模型)
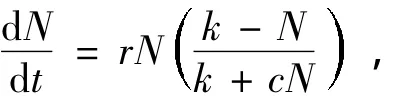
这里r,k,c是正常数,许多学者从不同角度对Smith模型及其各种推广形式进行了深入研究[9-11]。
据悉,文献中对于食饵种群遵循Smith增长和捕食者种群具有HollingⅡ型功能性反应的捕食者-食饵模型至今尚未有学者研究。为此,本文考虑如下自治的具有双密度制约捕食者-食饵系统

其中,x(t),y(t)分别表示时刻食饵和捕食者的种群密度,系数r,a,b,c,α,β,k,ω均为正常数。系统(1)的建立基于以下假设:
(1)食饵种群:食饵种群的相对增长率(即平均增长率)遵从Smith增长函数h(x)=(r-ax)/(1+kx),并且满足h(0)=r>0,h(B)=0,h'(x)=-(a-kr)/(1+kx)2<0,(0≤x<+∞,B=r/a)。
(2)捕食者种群:捕食者种群的功能性反应函数φ(x)=αx/(1+ωx)为HollingⅡ型,而β=ea,e表示转化系数,d为捕食者种群的死亡率,b为捕食者种群的密度制约系数。
(3)考虑到r,β分别为系统(1)中的食饵种群和捕食者种群的增长系数,本文总是假设β-dm>0,d/(β-dm)<B。
自治的模型(1)具有一般性,其特殊情形在文献中已被广泛讨论。如k=0,m=0即经典的Lotka-Volterra捕食者-食饵系统[12,13];又如k=0即通常称之谓Rosenzweig-MacArthur模型[14,15]。
1 平衡点的性态

则由
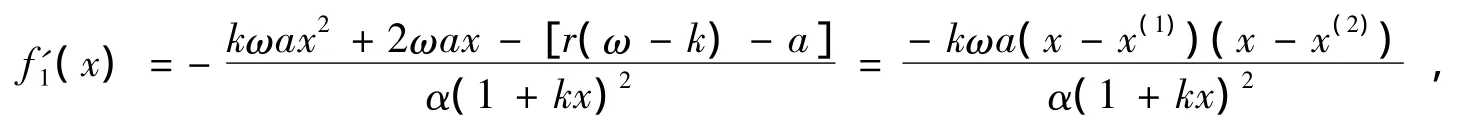
其中

可知x(1)>0,x(2)<0,当0<x<x(1)时(x)>0,当x>x(1)时(x)<0,得极大点x(1),记M=f1(x(1)),注意到

由x(1)<B知在x∈(0,x(1)),f1(x)是凸的上升;在x∈(x(1),B),f1(x)是凸的下降。
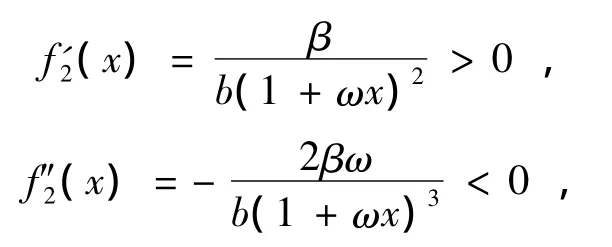

那么系统(1)存在唯一正平衡点Ω(x0,y0),其中αy0=(1+ωx0)h(x0),并且
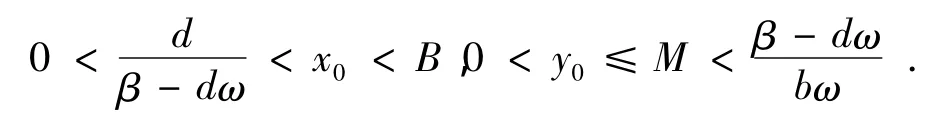
1)在O(0,0)处,系统(1)的线性化系统的系数矩阵为
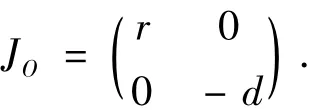
由于detJO=-rd<0,所以O为系统(1)的鞍点。
2)在E(B,0)处,系统(1)的线性化系统的系数矩阵为
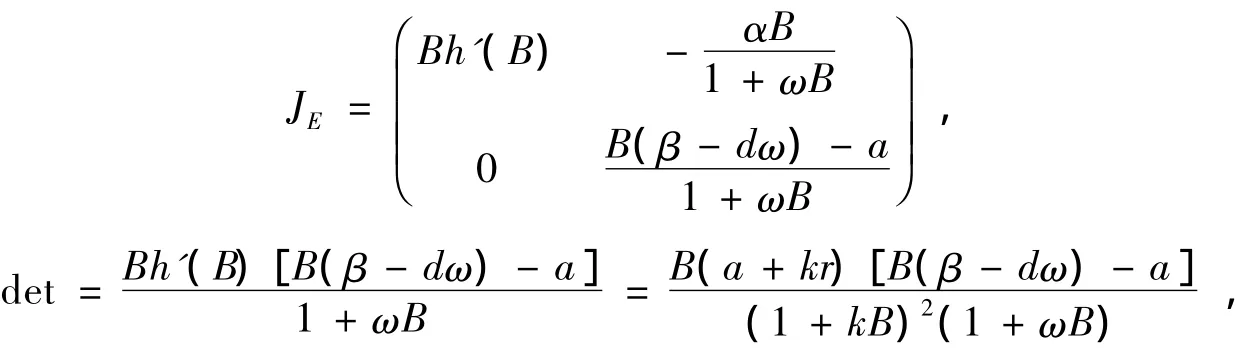
由于B>d/(β-dω),detJE<0,所以E是系统(1)在内的鞍点。
3)在Ω(x0,y0)处,系统(1)的线性化系统的系数矩阵为

这里,N=(1+ωx0)h'(x0)+ωh(x0),当r(ω-k)-a>aωx0(2+kx0)时,恒有N>0成立。注意到,因此

由此,可得如下结论:
①当r(ω-k)-a>aωx0(2+kx0)且可知Ω是系统(1)在内的稳定焦(结)点;
②当r(ω-k)-a>aωx0(2+kx0)且知Ω是系统(1)在内的不稳定焦(结)点;
③当r(ω-k)-a>aωx0(2+kx0)且,则由文[8]定理4.3知Ω是系统(1)在内的中心-焦点型奇点。
引理1非负平衡点O(0,0)和E(B,0)是系统(1)的鞍点,对于正平衡点Ω(x0,y0):
①当r(ω-k)-a>aωx0(2+kx0)且时,Ω是系统(1)的稳定焦(结)点;
②当r(ω-k)-a>aωx0(2+kx0)且时,Ω是系统(1)在的不稳定焦(结)点;
③当r(ω-k)-a>aωx0(2+kx0)且时,Ω是系统(1)在内的中心-焦点型奇点。
证明设x1=max{,B}作直线l1:x-x1=0,即x=x1,当x1=B时,由系统(1)得

当x1=时,表明>B(即r-a<0),由系统(1)得
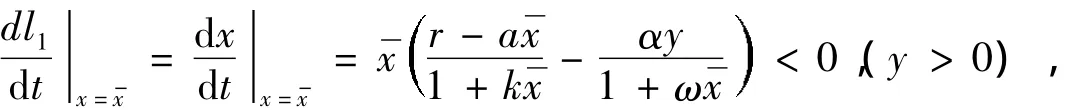
这表明系统(1)的正半轨线经直线l1时是自右向左的;

可见,系统(1)的正半轨线经直线l2时是自上向下的;
又直线l3:x-0=0及l4:y-0=0都是系统(1)的轨线,因此由l1,l2,l3,l4构成的闭合曲线Γ所围成的区域是系统(1)的最终有界区域。证毕。
2 闭轨线的不存在性
定理1如果2b<α≤4b且2b(rω+α)>αa,则系统(1)在内不存在闭轨线
证明作Dulac函数:B(x,y)=x-1y-1,于是,由系统(1)及∀(x,y)∈,有
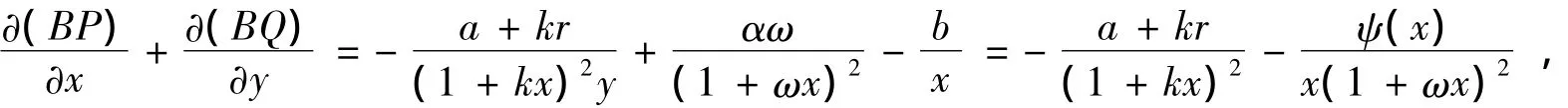
其中ψ(x)=b(1+ωx)2-αωx,由于

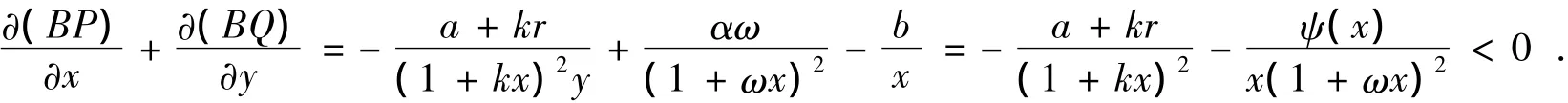
定理2设系统(1)满足定理1条件,又同时满足条件

证明由引理1、引理2和定理1易得,证略。
3 极限环的存在唯一性
首先,我们讨论系统(1)极限环的存在性:由引理1知,Ω(x0,y0)是系统(1)的不稳定焦(结)点。今作包围不稳定焦(结)点Ω(x0,y0)的闭区域:
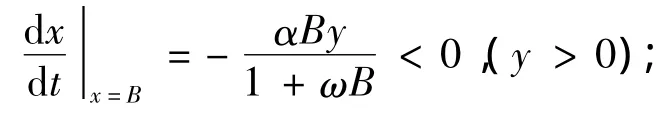
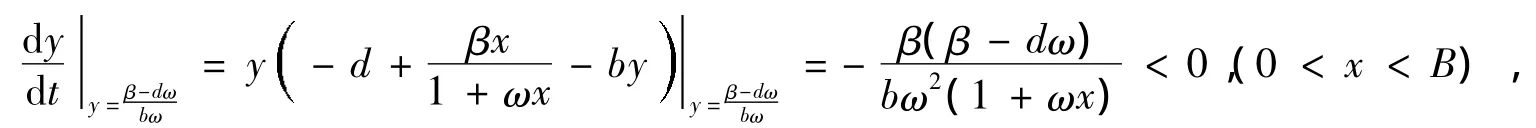
③直线x=0及y=0,即正半y轴和正半x轴都是系统(1)的轨线。
再证唯一性:作自变数变换:(1+ωx)dτ=dt,则将系统(1)化为下面等价系统

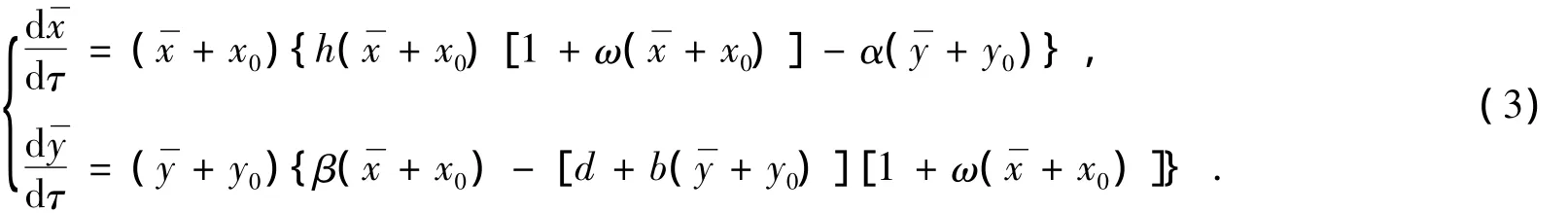


再令αy0exp{b[ωx0(eu-1)+u]/α}dτ=ds,变换后u,υ,ds仍记为x,y,dt,进而将系统(1)化为Lienard系统

其中
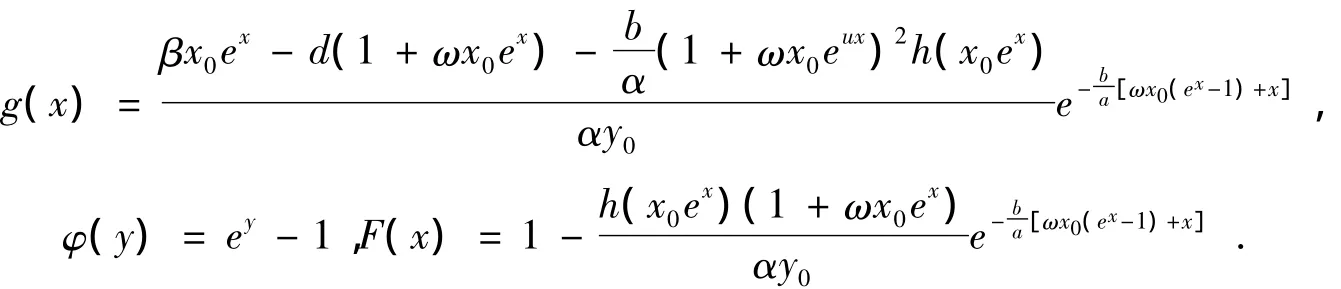
由上述所使用的一系列变换可知,系统(1)的正平衡点Ω(x0,y0)变为Lienard系统(6)的平衡点O(0,0),为此,相应地需要证明Lienard系统(6)在包含有平衡点O(0,0)的带状区域-∞<x<ln{B/x0}内,至多存在一个极限环。
由系统(1)在带状区域0<x<B内存在唯一正平衡点Ω(x0,y0)知
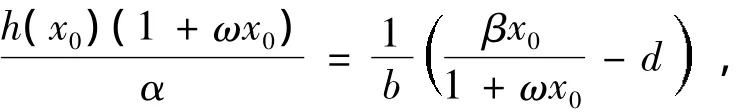
据此,H(x)=0存在唯一正根x0使得H(x0)=0,当0<x<x0时,即f2(x)<f1(x);当x0<x<B时,f2(x)>f1(x),即H(x)>0。
①由所使用变换知,当0<x<B时,对应系统(6)中-∞<x<ln{B/x0},当0<x<x0时,H(x)<0,ln{x/x0}<0,对应于-∞<x<0,xg(x)>0;当x0<c<B时,H(x)>0,ln{x/x0}>0,对应于0<x<ln{B/x0},xg(x)>0.因此,x≠0,xg(x)>0.
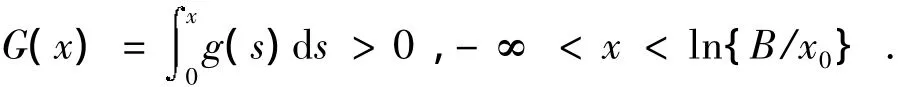
②y≠0,yφ(y)=y(ey-1)>0,φ(-∞)=-1,φ(+∞)=+∞,并且φ'(y)=ey>0,φ(y)是单调增加的。
③由前面讨论知αy0=(1+ωx0)h(x0),所以F(0)=1-1=0,在条件
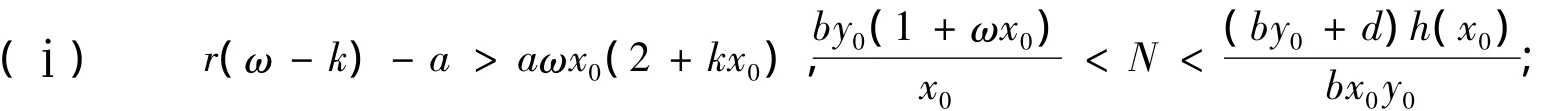
当-∞<x<ln{B/x0}且x≠0时,相应于系统(1)0<x<B且x≠x0,则当条件

成立时,恒有
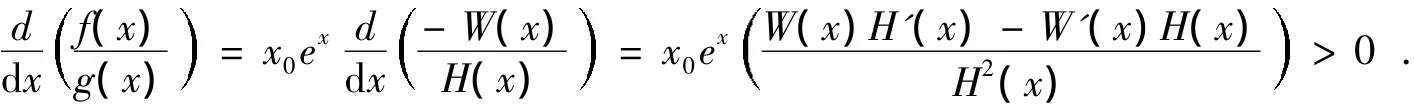
综上,根据张芷芬唯一性定理[16]获得如下定理3。
定理3如果系统(1)满足条件

6 结论
本文所研究的食饵带有Smith增长的捕食者——食饵模型,具有重要的理论意义和实际应用价值。当参数满足定理2条件时,捕食者——食饵两种群将长期共存并稳定在一组恒正的定值上;当参数满足定理3条件时,捕食者——食饵两种群将产生生物性周期振荡现象,并将趋于稳定的。这一结果不仅揭示了该生态系统在食物有限时的基本演变规律,而且在控制理论中也有其重要学术价值。
[1]Holling C S.The Functional Response Predators to Prey Density and its Role in Mimicty and Population Regulation[J].Mem Ent Sci Cannad,1965,97(45):5-60.
[2]匡亦群,邱梅青.一类具功能反应的食饵-捕食者模型的定性分析[J].生物数学学报,2007,22(4):629-633.
[3]Jicai Huang.Bifurcation and Chaos in a Discrete Predator-Prey System with Holling Type-ⅣFunctional Response[J].Acta Math.Appl.Sinica,English series,2005,21(1):157-176.
[4]田德生,朱长青,朱永松.HollingⅣ捕食-食饵时滞系统的多个周期解[J].纯粹数学与应用数学,2009,25(2):339-345.
[5]黄健民.食饵按广义生长的HollingⅡ型功能反应的捕食者-食饵模型[J].华南师大学报,1996(4):90-95.
[6]Fan M,Wang K.Global Existence of Positive Periodic Solution of a Predator-Prey System with Holling Type II Functional Response[J].Acta Mathematiea Scintia,2001,21(4):83-91.
[7]Li Y K.Periodic Solutions of a Periodic delay Predator-Prey System[J].Prie of Amer Math Sec,1992,127(5):1331-1335.
[8]Smith F E.Population Dynamics in Daphnia Magna and a new Model for Population Growth[J].Ecology,1963,44(4):651-663.
[9]孙伟平.微分方程边值问题的若干研究[D].北京:北京理工大学论文,2000:65-68.
[10]Lotka A J.Elements of Physical Biology,Williams and Wilking Baltimore[M].New York:Reissued as Elements of Mathematical Biology,Dover,1956.
[11]Volterra V.Variazienie Fluttuazini Del Numere in Specie Animali Conviventi[M].Mem Res Com Tolassogr Ital,1927.
[12]Lotka A J.Elements of Physical Biology,Williams and Wilking Baltimore[M].New York:Reissued as Elements of Mathematical Biology,Dover,1956.
[13]Rosengzweig M L.Why the Prey curve has a Hump[J].The American Naturalist,1969,103(929):81-87.
[14]陈兰荪,井竹君.捕食者-食饵相互作用微分方程的极限环存在唯一性[J].科学通报,1984,24(9):521-523.
[15]张芷芬,丁同仁,黄文灶.微分方程定性理论[M].北京:科学出版社,1985,152-153.
猜你喜欢
力学学报(2022年12期)2023-01-15
数学物理学报(2022年1期)2022-03-16
数学物理学报(2021年6期)2021-12-21
湖北工程学院学报(2021年3期)2021-06-16
宜宾学院学报(2020年12期)2021-01-30
应用数学(2020年4期)2020-12-28
数学物理学报(2019年6期)2020-01-13
数学物理学报(2019年4期)2019-10-10
数学物理学报(2019年3期)2019-07-23
数学物理学报(2018年5期)2018-11-16

- 东北电力大学学报的其它文章
- 烟塔合一技术对循环水水质的影响