
河南省主推小麦品种越冬期生理生化特性比较研究
2012-02-05 09:47张自阳冯素伟姜小苓李淦胡铁柱茹振钢
河南科技学院学报(自然科学版) 2012年4期
张自阳,冯素伟,姜小苓,李淦,胡铁柱,茹振钢
(河南科技学院,河南新乡453003)
河南省属暖温带向亚热带、湿润向半湿润过渡的季风气候区,冬季寒冷雨雪少,低温冻害是黄淮麦区小麦生产上的主要气象灾害之一.近年来河南省灾害性天气频繁,给农业生产造成巨大损失,特别是低温对河南省小麦生产构成巨大威胁.据2005年1月12日统计[1],小麦冻害面积143.79万hm2,严重冻害面积24.73万hm2;2007年3月7日、4月3日河南省出现两次强度较大的降温过程,造成大部分小麦品种受冻害,全省小麦受冻害面积达16.04万hm2,有0.67万hm2绝收,对小麦生产造成了巨大影响;2010年出现了零下16℃的极端低温等.小麦冻害已成为小麦生产发展亟待解决的重要问题,而选用抗冻性强的小麦品种是有效防止或减轻冻害损失的基本途径.
对于小麦的抗寒机理已有较多的研究[2-4],不同研究者对不同地区、不同抗寒品种的生理指标界定也不尽相同[5-6].国审小麦新品种百农矮抗58自审定以来经受了干旱、低温等多种不良天气的严峻考验,现在已成为推广面积最大、发展势头最好的全国主导品种.本试验对百农矮抗58和河南省主栽小麦品种的生理、生化特性进行比较研究,旨在为百农矮抗58的抗寒性研究、黄淮地区半冬性小麦品种抗寒鉴定及小麦新品系抗寒性评价提供生理生化方面的理论依据.
1 材料与方法
1.1 材料与试验设计
供试品种为百农矮抗58、周麦18、郑麦9023、豫麦49、郑麦7698、周麦22.试验材料均由河南科技学院小麦中心提供.
试验于2010-2011年在河南科技学院试验田进行.随机区组设计,3次重复,行长3 m,6行区,行距0.23 m,田间管理同一般高产田.
1.2 取样与测定方法
试验材料从2011年1月12日开始取样,气温变化见图1(取样前20 d至取样结束)所示.每隔10 d取样1次,共3次,每个品种随机选取长势一致的麦苗50株,用蒸馏水小心冲洗,晾干,样品在冰盒上分苗,剪取整个分蘖节和叶片,将叶片和分蘖节均剪成0.5 cm长的小段,混匀,按0.5 g分装,置-80℃冰箱贮存备用.考马斯亮蓝G-250法测定可溶性蛋白含量,酸性茚三酮法测定脯氨酸含量[7],蒽酮比色法测定可溶性糖含量[8],电导仪(DDS-307)测定相对电导率[9],愈创木酚法测定过氧化物酶(POD)活性[7],均3次重复.

图1 最高温度与最低温度变化图(2010/12/20-2011/02/08)Fig.1 The lowest temperature and the highest temperature during2010-12-20 to2011-02-08
1.3 数据处理
应用Microsoft Excel 2003,SAS9.0进行数据处理.
2 结果与分析
2.1 相对电导率的变化
河南省主推小麦品种越冬期相对电导率的变化见图2.

图2 不同小麦品种叶片相对电导率的变化Fig.2 The changes of relative electric conductivityin the leafofdifferent wheat varieties
由图2可知,6个小麦品种的叶片相对电导率均出现了先升高后降低的变化过程.这是由于小麦叶片在遭受低温影响时,细胞膜受到伤害,细胞内的各种水溶性物质包括电解质有不同程度的外渗,导致电导率上升,但随着低温对叶片的持续影响,小麦在经受了低温抗寒锻炼之后,通过自身的调节适应了外在环境的变化,使细胞膜得以修复,又出现了电导率降低的现象.
图2表明,1月15日与2月6日取样的6个小麦品种中,百农矮抗58的相对电导率均小于其他5个品种,1月26日取样的百农矮抗58与郑麦7698相对电导率均低于其他4个品种.说明在低温胁迫下百农矮抗58的原生质膜稳定性好,抗寒性相对较强.
2.2 可溶性糖含量的变化
河南省主推小麦品种越冬期可溶性糖含量的变化见表1.

表1 不同小麦品种各器官可溶性糖含量的变化Tab.1 Change ofsoluble sugar content in the organs of different wheat varietiesmg/g
由表1可知,在低温条件下,不同品种间可溶性糖在不同器官、不同取样时期差异均达到了显著水平,且分蘖节中的可溶性糖含量高于叶片中的含量.
2011年1 月15 日取样的百农矮抗58分蘖节中可溶性糖含量高于其他5个品种,并且达到了显著差异;1月26日取样的百农矮抗58分蘖节中可溶性糖含量高于其他5个参试品种,且显著高于郑麦9023;2月6日取样的分蘖节中,百农矮抗58的可溶性糖含量稍低于郑麦7698,但明显高于其他4个品种,显著高于郑麦9023和豫麦49.
由6个参试品种叶片可溶性糖含量检测中可以看出,1月15日取样的百农矮抗58叶片中可溶性糖含量稍低于周麦18,但高于其他4个品种,显著高于郑麦9023;1月26取样的叶片中,百农矮抗58可溶性糖含量最高,且显著高于郑麦7698;2月6日取样的叶片中,百农矮抗58可溶性糖含量均高于其他5个品种,且显著高于郑麦9023、周麦22、郑麦7698.
2.3 可溶性蛋白含量的变化
植株体内的可溶性蛋白质多为一些代谢酶类,其含量多少反映了植株活力的强弱.河南省主推小麦品种越冬期可溶性蛋白含量的变化见表2.

表2 不同小麦品种可溶性蛋白含量的变化Tab.2 Changes of soluble protein content in the organs of different wheat varietiesmg/g
由表2可知,6个参试品种叶片中的可溶性蛋白含量均高于分蘖节,且可溶性蛋白含量在品种间差异达到显著水平.
百农矮抗58在3个取样时期的叶片中可溶性蛋白含量均有较多的积累,1月15日取样的叶片中百农矮抗58的含量高于其他5个品种,且显著高于周麦18;1月26日叶片取样中,可溶性蛋白含量稍低于豫麦49,两者含量差异不显著,但高于其他4个品种,且可溶性蛋白含量差异与郑麦9023、周麦18、周麦22达到显著水平;2月6日取样的叶片中百农矮抗58的可溶性蛋白含量高于其他5个品种.同样,百农矮抗58的可溶性蛋白含量在分蘖节中依然较高.
以上结果表明,百农矮抗58可溶性蛋白含量在3个时期整体比较高,说明百农矮抗58具有较强的氮代谢功能.
2.4 脯氨酸含量的变化
河南省主推小麦品种越冬期脯氨酸含量的变化见表3.
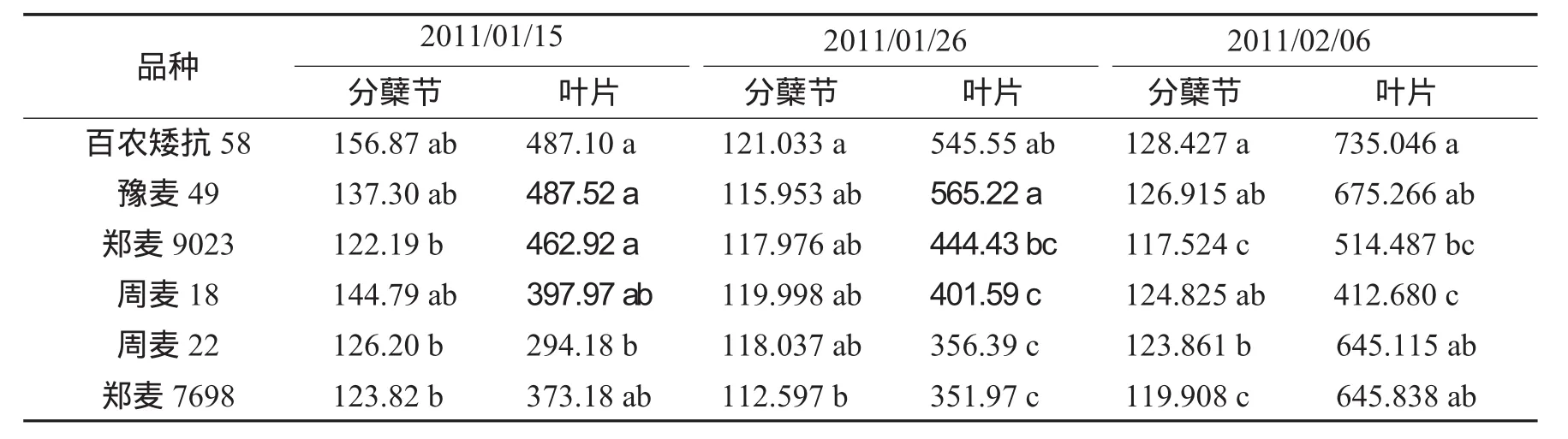
表3 不同小麦品种脯氨酸含量的变化Tab.3 Changes ofproline content in the organs of different wheat varietiesμg/g
由表3可知,在低温胁迫下各参试品种叶片中游离脯氨酸含量处于较高水平,并且在3个取样时期呈递增趋势.分蘖节中脯氨酸含量没有明显的递增趋势.6个品种3个取样时期叶片中的游离脯氨酸含量均高于分蘖节.不同品种、不同取样时期脯氨酸含量差异达到了显著水平.
1月15 日取样的分蘖节中,百农矮抗58脯氨酸含量高于其他5个品种,且显著高于郑麦9023、周麦22、郑麦7698;1月26日取样的分蘖节中,百农矮抗58分蘖节中脯氨酸含量均高于其他5个品种,显著高于郑麦7698;2月6日取样分蘖节中脯氨酸含量最高,且显著高于郑麦9023、周麦22、郑麦7698.
从6个参试品种叶片中的游离脯氨酸含量变化可以看出,1月15日取样的叶片中,百农矮抗58的脯氨酸含量低于豫麦49,但高于其他4个品种;1月26日取样的叶片中百农矮抗58的脯氨酸含量显著高于周麦18、周麦22、郑麦7698;2月6日取样的叶片中,百农矮抗58脯氨酸含量最高,显著高于郑麦9023、周麦22.
2.5 POD活性的变化
河南省主推小麦品种越冬期分蘖节中POD活性的变化见图3.

图3 不同小麦品种分蘖节POD活性变化Fig.3 Changes of PODactivity in the tilleringnode of different wheat varieties
由图3可知,在低温持续影响下,6个品种的POD活性在分蘖节中均呈现逐渐增高的趋势.1月15日与1月26日取材的分蘖节中,百农矮抗58的POD活性高于其他5个品种;2月6日取材的分蘖节POD活性稍低于周麦22,高于其他4个品种.
河南省主推小麦品种越冬期分蘖节中POD活性的变化见图4.

图4 不同小麦品种叶片中POD活性变化Fig.4 Changes of PODactivityin the leaves of different wheat varieties
由图4可知,在低温持续影响下,除豫麦49外,其余5个品种的POD活性在叶片中均呈现逐渐增高的趋势.1月15日、1月26日取样的百农矮抗POD活性稍低于周麦18,但均高于其他4个品种;2月6日取样的百农矮抗58的POD活性高于其他5个品种.
3 结论与讨论
植物的抗寒性受基因型的控制,同时,与其自身代谢和生理过程关系密切.植物在低温逆境下生长发育受阻,诱发植物体内的抗冻基因合成大量的蛋白以及抗逆境的小分子物质[10].例如,诱导蛋白质的合成、可溶性糖的积累,质膜透性发生改变,抗氧化物及多种代谢酶增加等[11].
本试验结果表明:在低温胁迫下,与其他5个品种相比,百农矮抗58分蘖节和叶片中可溶性糖、可溶性蛋白、脯氨酸含量整体较高.并且随着低温的持续作用,脯氨酸含量呈明显的递增趋势,电导率变化规律比较明显,可溶性糖和可溶性蛋白没有明显的增加趋势,但都维持在一个较高的水平.
低温胁迫下可溶性糖在植物体内逐渐积累,可溶性糖含量较高的植物细胞的溶质浓度增大,缓和胞外结冰引起的细胞失水,增强质膜的稳定性[12],有利于抗寒力的提高.许多研究证明,小麦抗寒性与植株糖含量存在正相关的关系,是衡量小麦抗寒性的一个重要指标[5,13-14].对百农矮抗58叶片和分蘖节的研究结果表明:在持续低温的影响下,其分蘖节和叶片中积累了大量的可溶性糖,分蘖节中可溶性糖含量大于叶片,说明分蘖节的抗寒适应能力强.含糖量高可以改变细胞溶质的浓度,增强质膜的稳定性,这一点在对其电导率的研究结果中得到证实,百农矮抗58三个取样时期电导率低于其他5个品种,说明其质膜透性比较稳定,矮抗58的抗寒能力较强.可以把小麦叶片和分蘖节中可溶性糖含量和相对电导率作为半冬性小麦品种抗寒性鉴定的一个重要指标,这与高志强等研究结果相似[5].
许多学者认为脯氨酸含量与高等植物的抗寒性存在相关性,脯氨酸在体内调节渗透压[15]、降低水势、解毒并降低冰点[16].对百农矮抗58的研究中发现,在低温持续影响下,其体内脯氨酸含量较高,随着低温时间延长脯氨酸含量呈递增趋势.与其他5个品种相比较,百农矮抗58体内脯氨酸含量整体上较高.叶片中的脯氨酸含量有一个比较明显的递增趋势.
POD是植物细胞的保护酶,POD活性越高,其抗寒能力越强.对百农矮抗58 POD活性的研究结果表明:分蘖节和叶片中的POD随低温胁迫时间的增长呈明显的递增趋势,与百农矮抗58在2月6日取样电导率变小有一定的关系,说明此期间分蘖节和叶片的清除自由基的能力强,质膜过氧化程度轻,电解质外渗较少导致电导率较小.
从以上分析中得出如下结论:①供试的6个小麦品种中,百农矮抗58的抗寒性相对较强;②低温胁迫下叶片相对电导率、分蘖节可溶性糖含量、叶片脯氨酸含量变化规律较明显,可以作为半冬性品种和新品系抗寒性辅助鉴定的重要指标.
[1] 陈咏雪.河南小麦生产中灾害性天气的应变栽培技术[J].河南农业,2009(11):51.
[2] 王多佳,曾俨,牟永潮.高寒地区冬小麦东农冬麦1号抗冻蛋白的研究[J].麦类作物学报,2009,29(5):823-826.
[3] 于晶,张林,崔红,等.高寒地区冬小麦东农冬麦1号越冬前的生理生化特性[J].作物学报,2008,34(11):2019-2025.
[4] 王红星,吴诗光,杨光宇.寒胁迫对小麦幼苗生理生化特性的影响[J].周口师范高等专科学校学报,2000,17(2):4-5.
[5] 高志强,张国红,张爱芝,等.不同小麦品种对低温的生理反应研究[J].陕西农业大学学报,2002,22(2):109-112.
[6] 孙金月,赵玉田,梁博文,等.HRGP在小麦抗寒锻炼过程中的变化及其与抗寒性的关系[J].植物遗传资源学报,2004,5(1):6-11.
[7] 郝再彬,苍晶,徐仲.植物生理实验[M].哈尔滨:哈尔滨工业大学出版社,2004:46-49.
[8] 王学奎.植物生理生化实验原理和技术[M].北京:高等教育出版社,2006:202-204.
[9] 中国科学院上海植物生理研究所,上海市植物生理学会.现代植物生理学实验指南[M].北京:科学出版社,1999.
[10] Mastrangelo A M,Belloni S,Barilli S,et al.Low temperature promotes in tron retention in two e-cor genes of durum wheat[J].Planta,2005,221(5):705-715.
[11] Minami A,Nagao M,Ikegami K,et al.Cold acclimation in bryophytes:low-temperature-induced freezing tolerance in Physcomitrella patens is associated with increases in expression levels of stress-related genes but not with increase in level of endogenous abscisic acid[J].Planta,2005,220(4):414-423.
[12] Galiba G,Kerepesi I,Vagujfalvi A,et al.Mapping of genes involved in glutathione,carbohydrate and COR14b cold induced protein accumulation duringcold hardeningin wheat[J].Euphytica,2001,119(1/2):173-177.
[13] 杨春玲,宋志均,陈玉香,等.小麦抗寒机理研究[J].山西农业科学,2008,36(1):49-52.
[14] 于晶,张林,苍晶,等.冬小麦抗寒性的研究进展[J].东北农业大学学报,2008,39(11):123-127.
[15] 李志,王刚,吴忠义,等.脯氨酸与植物抗渗透胁迫基因工程改良研究进展[J].河北师范大学学报:自然科学版,2005,29(4):405-408.
[16] 汤章城.逆境条件下植物脯氨酸的积累及可能的意义[J].植物生理学报,1984,10(1):15-20.
猜你喜欢
中老年保健(2022年3期)2022-11-21
作物学报(2022年9期)2022-07-18
耕作与栽培(2022年1期)2022-04-07
麦类作物学报(2022年1期)2022-03-01
粮食加工(2020年4期)2020-02-28
农民致富之友(2019年33期)2019-12-20
科学与财富(2019年15期)2019-10-21
中国瓜菜(2019年4期)2019-07-03
发酵科技通讯(2018年2期)2018-07-06
