
长白楤木茎结构观察
2011-12-28 02:14:24樊清伟刘秀岩朱俊义
通化师范学院学报 2011年12期
樊清伟,杨 东,刘秀岩,朱俊义
(1.通化市城市绿化管理处,吉林通化134002;2.通化师范学院,吉林 通化134002)
长白楤木茎结构观察
樊清伟1,杨 东1,刘秀岩2,朱俊义2
(1.通化市城市绿化管理处,吉林通化134002;2.通化师范学院,吉林 通化134002)
利用扫描电子显微镜技术,观察长白楤木(Aralia contunentalisKitag)茎结构,研究结果是:初生维管束有两种排列方式同时存在,即一种是排列成一轮在皮层薄壁组织的外侧,一种是散生在皮层薄壁组织中,由木质部和韧皮部组成.次生维管束位于成轮排列的维管束之间,由进行次生生长时由两初生维管束之间的束间形成层形成.
长白楤木;茎;次生结构
长白楤木(Aralia contunentalisKitag)属于五加科楤木属多年生草本植物[1].春季长出的长白楤木嫩芽是东北地区传统山野菜,具有独特的清香味道,介于刺嫩芽与香椿之间,王忠壮等(1999)对楤木属药食兼用嫩芽的营养成分进行研究的结果表明,长白楤木有很高的营养价值,其吃法很多,可直接炒食、生拌、蘸酱等,也可盐渍[1-3].本文研究其茎结构,对指导其反季栽培管理有重要意义.
1 实验材料与方法
1.1 实验材料
取材地点:通化师范学院院内生物专业试验基地栽培的长白楤木,海拔400m.茎用FAA固定液长期固定备用.
1.2 BSE2观察模式的电镜观察标本的制作
长白楤木茎结构可通过扫描电镜在BSE2观察模式下观察.取固定的实验材料,用清水洗掉FAA固定液,首先用无水乙醇连续脱水3次,每次24h,再用二甲苯连续浸泡2次,每次24h,之后再用无水乙醇连续浸泡3次,每次24h,以溶解掉材料内的二甲苯,最后将材料放入清水中,以待解剖观察.将处理好的实验材料置体视镜下解剖,挑选观察的内容放在样品台导电胶上,置HITACHI S-3000N型扫描电子显微镜内,在低真空背散射模式(BSE2)下观察照相.
2 实验结果
长白楤木茎横切观察可见,其茎由表皮、皮层厚角组织、皮层薄壁组织、初生维管束、次生维管束组成.初生维管束有两种排列方式,一种是排列成一轮在皮层薄壁组织的外侧,一种是散生在皮层薄壁组织中,由木质部和韧皮部组成.次生维管束位于成轮排列的维管束之间,是进行次生生长时由两初生维管束之间的束间形成层形成的,束中形成层活动较弱.从茎的纵切看,导管分子壁具梯纹、孔纹增厚.
3 讨论
植物茎结构中维管束排列主要有多种方式,即双子叶植物木本植物维管束在皮层中排列成一轮,维管束数量较多,排列紧密,在两个初生维管束之间形成次生维管束,在初生维管束的内部形成次生维管组织,进入次生生长,如短梗五加茎结构;双子叶植物草本植物维管束在皮层中排列成一轮,维管束数量较木本植物少,排列不紧密,在两个初生维管束之间形成小的次生维管束,进人次生生长,如向日葵茎结构,但多数双子叶植物草本植物维管束束间形成层活动弱,不进行次生生长;单子叶植物维管束在皮层中排列成两轮,如水稻、小麦,或散生在基本组织当中,如玉米、高粱.多数单子叶植物茎维管束束间形成层活动弱,不进行次生生长[4-6].
五加科植物茎有草本和木本之分,内部结构较为复杂.长白楤木为草本植物,本文研究结果表明长白楤木茎由表皮、皮层厚角组织、皮层薄壁组织、初生维管束、次生维管束组成.初生维管束有两种排列方式同时存在,即一种是排列成一轮在皮层薄壁组织的外侧,一种是散生在皮层薄壁组织中,由木质部和韧皮部组成.次生维管束位于成轮排列的维管束之间,是进行次生生长时由两初生维管束之间的束间形成层形成的,束中形成层活动较弱.长白楤木维管束的两种排列方式即散生、排列成轮同时存在的情况未见报道,其同时具有双子叶植物草本植物和单子叶植物茎维管束的排列特征,并且,维管束数量较多,说明长白楤木茎结构比较复杂,介于木本植物和草本植物之间.在栽培过程中,应加强水、肥、光照的管理,减少其次生长,生产高质量的嫩茎产品.
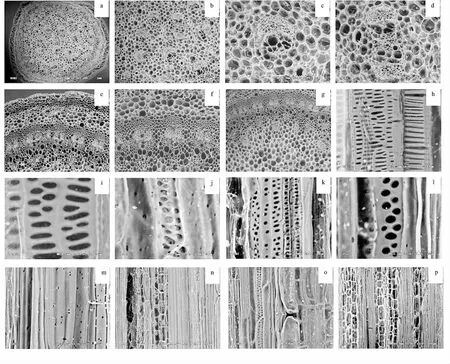
图1 长白楤木茎结构
[1]何景,曾沧江.中国植物志[M].第五十四卷.北京:科学出版社,1978.
[2]王忠壮,靳守东,全山丛,等.楤木属植物药食兼用嫩芽营养成分分析[J].营养学报,1999,21(1):100-103.
[3]姜晓莉.刺嫩芽山野菜的开发利用[J].农牧产品开发,1999(7):13-14.
[4]谷安根,陆静梅,王立军.维管植物演化形态学[M].长春:吉林科学技术出版社,1993.
[5]陆时万,徐祥生,沈敏健.植物学[M].北京:高等教育出版社,1982.
[6]A.FAHN.植物解剖学[M].吴树明,译.天津:南开大学出版社,1990.
Q944
A
1008-7974(2011)12-0034-02
吉林省科技发展计划项目(20111706).
2011-09-25
樊清伟(1970-),男,吉林通化人,通化市城市绿化管理处工程师.
(责任编辑:陈衍峰)
猜你喜欢
现代交际(2022年10期)2022-11-01 08:06:02
当代音乐(2022年8期)2022-04-29 16:02:48
河北农业大学学报(2021年5期)2021-11-10 13:11:28
林业科学(2020年10期)2020-11-30 00:36:58
西藏人文地理(2020年4期)2020-11-19 04:32:00
新疆农业科学(2020年9期)2020-10-13 12:37:42
农业工程技术·综合版(2020年1期)2020-04-23 09:38:30
当代音乐(2019年11期)2019-11-11 02:23:50
河南农业科学(2019年9期)2019-09-24 09:10:48
山东林业科技(2019年2期)2019-06-03 10:10:54
