
转基因棉花的研发现状与发展策略
2011-11-30 03:12刘晨曦吴孔明
植物保护 2011年6期
刘晨曦, 吴孔明
(植物病虫害生物学国家重点实验室,中国农业科学院植物保护研究所,北京 100193)
棉花(Gossypium hirsutum L.)隶属于锦葵科(Mal vaceae)棉属(Gossy piu m),是世界上最主要的经济作物之一。全世界植棉国家和地区有96个,其中产量较高的国家有中国、美国、印度等。我国是世界植棉大国,总产量居世界第一,植棉业是农业的支柱性产业,对国民经济的发展至关重要。
虫害和草害对植棉业有很大的影响,制约了棉花的生产。我国棉花害虫多达300余种,时常大面积暴发成灾,造成棉花产量的严重损失。由于大量使用化学农药导致了一些棉花害虫抗药性的产生,严重威胁棉花生产,同时也使环境污染日益恶化。棉田杂草与棉花之间存在水分、阳光、空间和营养物质的竞争,严重影响了棉花的正常生长。为了减轻除草劳动强度、提高生产效率,化学除草剂应运而生。草甘膦等非选择性除草剂具有高效、低毒、易降解和无残留的优点,但由于对作物同样具有杀灭作用,而不能在作物的生长期应用。
利用植物基因工程和遗传育种技术手段培育的转基因抗虫、耐除草剂棉花,为棉花害虫和草害的控制提供了新的手段。转基因棉花自1996年商业化种植以来,有效控制了棉花害虫的为害,减少了化学杀虫剂的使用[1],降低了除草剂对棉花植株的损伤,扩大了除草剂的应用范畴。本文综述了国内外转基因棉花研发现状及其商业化进程,并针对其应用过程中存在的潜在风险,提出了我国转基因棉花应采取的研发策略。
1 转基因棉花研发动态
目前,转基因棉花的研发与培育已经在多个国家进行,其过程涉及基因克隆、遗传转化、性状鉴定、株系筛选、品种培育和安全性评价等多个环节,研发核心是筛选出结构完整、表达良好的株系,构建具有商业推广价值的“转化事件”。国内外对转基因棉花的研究包括抗虫、耐除草剂、抗病、纤维改良、抗旱和耐盐碱等[2-6],但研制成功和取得重要进展的主要为抗虫、耐除草剂和纤维改良性状。表1为近年来各国研发注册的抗虫(IR)、耐除草剂(HT)或复合性状(IR+HT)的转基因棉花转化事件[7]。

表1 转基因棉花转化事件
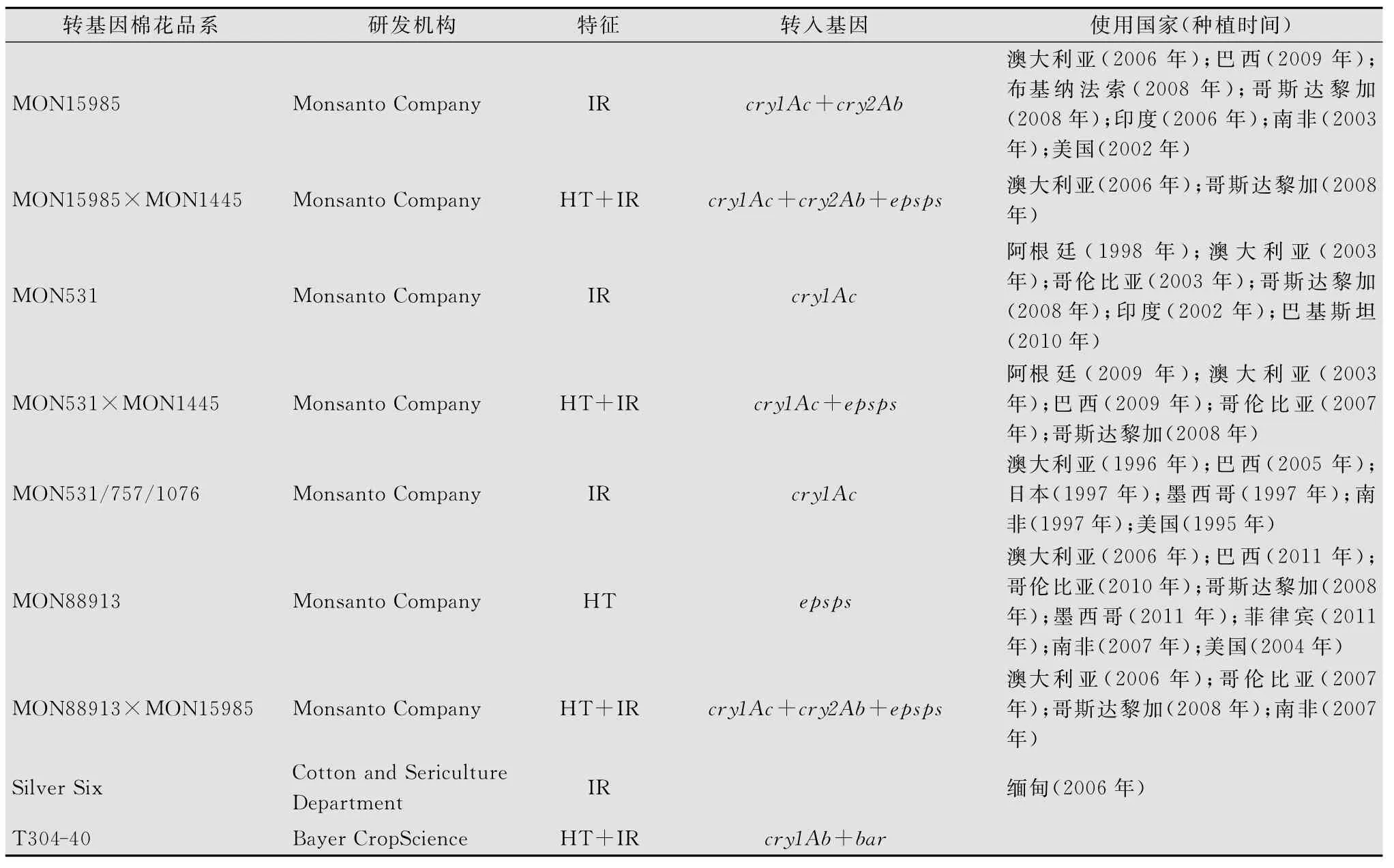
续表1
1.1 转基因抗虫棉花
1.1.1 防治鳞翅目害虫的Bt棉花
苏云金芽胞杆菌(Bacill us thuringiensis,Bt)在芽胞形成阶段会产生伴胞晶体,该晶体被鳞翅目昆虫取食后,在其中肠蛋白酶的水解作用下被激活,活化的毒素能与中肠特异性受体结合,导致中肠孔洞形成而使昆虫死亡[8]。目前发现并登记的cry1~cry67基因大约有190多种,其中cry1Ab基因、cry1Ac基因、cry1F基因、cry2Ab基因、cry2Ae基因等被用于转基因抗虫棉工程[9]。
1.1.2 防治棉盲蝽的Bt棉花
Bt棉表达的Cry杀虫基因对鳞翅目和鞘翅目昆虫有较好的防治效果,对半翅目的防效甚微。研究表明,长期种植Bt棉花引起的害虫地位演化导致棉田的次要害虫棉盲蝽发生数量增多,成为了Bt棉的主要害虫[10-12]。国外公司已针对这一问题开展了抗棉盲蝽转基因棉花研究,但尚未注册。如:Baum等[13]发现了一种新型的对棉盲蝽有较好防治效果的Bt杀虫蛋白基因,在转基因抗虫棉上表达的这种35 k D的Bt杀虫蛋白能够有效控制棉盲蝽的生长发育。
1.1.3 营养杀虫蛋白棉花
在Bt毒素研究过程中发现,一种在芽胞形成前的营养阶段分泌和产生另一种非δ-内毒素的杀虫营养蛋白,即Vip蛋白(vegetative insecticidal protein,Vip),被称为第2代杀虫蛋白。Vip蛋白与Bt Cry毒素的结构和功能完全不同,对鳞翅目和鞘翅目有很好的效果[14]。Syngenta公司(Greensboro,NC,USA)对表达Vip蛋白的转基因抗虫棉进行了多年的田间杀虫效果评估,结果表明该转基因抗虫棉能较好控制棉铃虫的种群[15]。
1.1.4 RNAi转基因棉花
植物介导的昆虫RNAi技术,可以有效、特异地抑制昆虫基因的表达,从而抑制害虫的生长,为农业害虫的防治提供了特异性更强且环境安全的新思路[16-21]。目前虽然国内外尚没有RNAi转基因抗虫棉花注册,但已成为十分活跃的研发领域。以RNAi技术为基础的转基因抗虫棉有望在将来的害虫防治领域中发挥重要作用。
1.2 耐除草剂棉花
1.2.1 耐草甘膦棉花
耐草甘膦基因是属于表达靶标酶类耐除草剂基因棉花。此类型的耐除草剂基因棉花产生靶标酶或靶标蛋白质,使棉花吸收除草剂后,仍然能进行正常代谢作用。5-烯醇式丙酮酰莽草酸-3-磷酸合成酶(EPSPS)是棉花氨基酸循环莽草酸代谢途径中一个很重要的酶。草甘膦通过抑制EPSPS,造成植物体内氨基酸代谢莽草酸途径的中断[22]。通过农杆菌介导法等方法,导入epsps基因的棉花植株对草甘膦有较好的耐性[22-23]。
1.2.2 耐草铵膦、双丙氨膦棉花
耐草铵膦、双丙氨膦棉花是属于表达修饰酶类的转基因耐除草剂棉花。此类型的耐除草剂棉花产生能修饰除草剂的酶或酶系统,在除草剂发生作用前对其进行修饰从而失去除草能力。bar基因编码乙酰转移酶基因(PAT),对草铵膦、双丙氨膦进行修饰,反应产物能被顺利代谢使其失去除草能力[24-25]。通过转入bar基因使棉花对草铵膦、双丙氨膦都有较好的耐性[25-26]。
1.2.3 耐磺酰脲类 (绿磺隆等)棉花
这类是属于表达异构酶类耐除草剂棉花,可以产生对除草剂不敏感的靶标酶或蛋白的异构体。surb-Hr a基因通过编码改变空间构型的乙酰乳酸合酶 (ALS)使磺酰脲类除草剂失去作用,从而保护植物的正常生理活动。同样psb A基因通过表达突变的光系统IIQB蛋白使植物体具有三氮苯类除草剂的耐性[27]。
1.3 纤维品质改良转基因棉花
棉花纤维是由胚珠外珠被表皮层的单细胞分化而来,其长度、强度、细度等品质性状是评价棉花质量的重要标准。提高棉纤维品质尤其是纤维强度,是当前棉花育种工作中的一个重点。传统育种在棉纤维品质改良方面起到了重要作用,但由于受到育种周期长、外源种质利用困难、产量较低等因素的限制,用常规育种技术进行纤维品质改良进展缓慢[28]。利用基因工程来取得这方面的突破值得探索。近年来,国内外科研人员相继开展了用外源纤维改良基因改良棉纤维品质的探索性研究并取得较快的进展。Zhang等[29]利用植物基因工程技术,通过外源启动子提高吲哚乙酸合成基因iaa M表达,从而促进了生长素在棉花胚珠上皮中的合成,棉纤维品质得到了较大的提高。
2 商业化现状
经过1986-1995年期间的室内研究和田间小规模的试验,在1996年,美国、澳大利亚等国家已经开始商业化种植转基因棉花,主要是抗虫和耐除草剂为主的转基因棉花。2010年,全球转基因棉花的种植面积达到2 100万h m2,其中转基因抗虫棉花(Bt棉花)的种植面积是1 610万hm2,转基因耐除草剂棉花(HT棉花)的种植面积是140万h m2,含有复合性状的转基因棉花(含Bt杀虫基因和耐除草剂基因)种植面积是350万h m2[7]。
2.1 中国
我国于1997年开始商业化种植Bt棉。Monsanto公司的转基因抗虫棉33B(cry1Ac)首先在河北省种植[2]。我国研发的Bt棉GK系列(改造的融合基因cry1Ab/cry1Ac)首先在安徽、山东、山西、湖北4个省种植[7]。1999年,中国开始商业化种植双价抗虫棉(cry1Ac+cpTI)[30]。2010年,转基因抗虫棉在中国的种植面积达到345万hm2,占棉花总种植面积的69%[7]。我国近年来对转基因耐除草剂棉花进行了研发与培育,但还处于田间试验阶段。
2.2 印度
农业在印度的国民经济中占有较大的比重,是其支柱产业。2010年,印度的棉花种植面积达到1 100万hm2,是世界上植棉面积最大的国家。Bt棉在印度的商业化种植始于2002年,种植面积大约5万hm2。2003年,种植面积翻了一番,在随后的几年中,种植面积不断扩大。2010年,Bt棉的种植面积为940万hm2,占总植棉面积的86%[7]。在印度,首先商业化种植的是单价抗虫棉Bollgard®Ⅰ(cry1Ac)。2006年,双价抗虫棉Bollgard®Ⅱ(cry1Ac/cry2Ab)开始种植,种植面积不断扩大,而从2007年起,单价棉的种植面积在逐年减少。2010年,双价棉的种植面积是660万h m2,而单价棉的种植面积减少到280万h m2[7]。据不完全统计,2011年,双价棉的种植面积将占Bt棉的90%[7]。2010年,印度的转基因棉花转化事件共有4个,这些棉花转化事件都是抗虫或耐除草剂为特征的转基因棉花,其中MON15985×MON88913是抗虫或耐除草剂为特征的转化事件,2mepsps是以耐除草剂为特征的转化事件,这些已在印度进入田间试验阶段,有望在2011-2015年进行商业化种植[7]。
2.3 美国
棉花是美国第五大作物,种植面积仅少于小麦,主要种植在南部和西部。美国主要种植两种转基因抗虫棉,一种是Monsanto公司研发的单价抗虫棉Boll gar d®Ⅰ,另一种是复合性状的转基因棉花。1996年,美国开始商业化种植转基因棉花,其中转基因抗虫棉种植面积达到73万hm2,到2001年,转基因抗虫棉的种植面积达到200万hm2。与2009年相比,在2010年转基因棉花的种植面积已经从320万h m2增加到390万hm2,其中Bt棉占15%,而(HT+Bt)棉占58%,HT棉占20%,而2011年,Bt棉占17%,HT棉占15%,(HT+Bt)棉的种植面积与2010年相比没有变化。(数据来源:美国农业部国家农业统计服务中心(NASS))。
2.4 澳大利亚
澳大利亚是世界第三大原棉出口国,主要种植在南部的新南威尔士州和北部的昆士兰州。澳大利亚于1996年开始种植转基因抗虫棉Bollgar d®Ⅰ,种植面积达3万h m2,以后种植面积逐年递增,至2000年已达16.5万h m2,而2001年,种植面积略有所下降(14.6万h m2)。在2002年和2003年两年期间,单价抗虫棉Bollgard®Ⅰ已经基本被双价抗虫棉Bollgard®Ⅱ替代。2007年,由于干旱使澳大利亚转基因棉花的种植面积大大降低。在2010年,大约91%的种植面积是复合性状(HT+Bt)的转基因棉花,而双价抗虫棉Bollgard®Ⅱ只占1%,转基因耐除草剂棉花(HT)占7%左右[7]。
2.5 巴基斯坦
位于南亚中部的巴基斯坦是一个以农业为主的国家,棉花是该国的主要经济作物。棉花生产位居世界第四。在2010年,巴基斯坦开始商业化种植转基因抗虫棉(8个常规品种和1个杂交品种),种植面积达240万h m2,占棉花总种植面积的75%。目前,巴基斯坦已成为第12个种植Bt棉的国家[7]。
3 我国发展对策
传统上我国棉花种植区主要分布于长江流域、黄河流域和西北内陆地区,随着产业结构的调整,新疆棉花种植面积逐年增加,已上升成为我国最大产棉区。总体上,我国棉花产业发展的长期性制约因素集中于:(1)长江流域、黄河流域棉铃虫和棉盲蝽的危害,新疆地区棉蚜等害虫的危害;(2)新疆地区和黄河流域棉花枯黄萎病的危害;(3)棉田杂草的危害;(4)棉花纤维品质不能满足纺织工业需求。针对不同棉花种植区的需求,培育抗病虫、耐除草剂和高品质复合性状转基因棉花对推动我国棉花产业发展有重要意义。
3.1 新型转基因棉花的研发
3.1.1 抗虫性状
我国于1997年开始商业化种植Bt棉,有效地控制了棉铃虫的种群,增加了有益天敌的数量,减少了农药的使用,取得了显著的经济、生态和社会效益。然而,随着Bt棉的广泛应用,关于害虫对Bt棉的敏感性降低的报道也逐年增多[31-32],表明害虫对转基因抗虫棉存在抗性风险[30,33]。同时,棉盲蝽种群由次要害虫上升为主要害虫或成为新的问题[10-12]。
转基因双价基因策略是指在同一转基因作物中表达两种不同杀虫机理的杀虫基因,这种策略可有效延缓抗性的进化[34]。双价转基因抗虫棉已经在印度、澳大利亚等国家广泛应用,有效地延缓了害虫对转基因抗虫棉的抗性。Zhang等[32]对大部分种植Bt棉的华北地区和有限种植Bt棉的西北地区的棉铃虫对Cry1 Ac毒素和Cr y2 Ab毒素的敏感性进行了分析,结果表明在华北的13个地区的棉铃虫种群对Cr y1 Ac毒素敏感性低于西北地区的2个种群,而对Cry2 Ab毒素的敏感性没有显著性差异。Gao等[35]对安次和夏津两地的棉铃虫种群对Cr y1 Ac毒素和Cr y2 Ab毒素的交互抗性进行了分析,结果表明这两个种群对这两种毒素均无交互抗性。An等[36]分析了安次和夏津两地的棉铃虫种群对Cry1 Ac毒素和Vip3 A毒素的敏感性,结果表明两个种群对Cr y1 Ac毒素和Vip3 A毒素的敏感性呈负相关。因此,双价转基因抗虫棉的研发和应用将会延缓棉铃虫种群抗性的演化。在我国Bt棉商业化种植后,棉田化学防治模式的改变等相关因素使棉田的次要害虫棉盲蝽上升为棉田主要害虫[10-12],需要开发防治棉盲蝽的转基因抗虫棉花[13,37]。此外,研发抗棉蚜棉花对新疆地区棉花害虫的防治有应用价值。
3.1.2 抗病性状
棉花枯萎病和黄萎病对棉花生产有着严重的影响,目前国内外的研发尚未取得较大的进展。因此,需要挖掘抗病功能基因研发转基因抗病棉花。
3.1.3 耐除草剂性状
我国还没有培育出转基因耐除草剂棉花品种,研发还处于早期阶段。应针对我国棉花的种植情况及杂草种群的类型,借鉴国外的转基因耐除草剂棉花的研发和使用经验,加快速度研发耐草甘膦等高效、低毒、低残留除草剂的转基因棉花,为我国转基因耐除草剂棉花的商业化种植奠定基础。
3.1.4 纤维品质改良性状
以棉纤维形成和发育的分子机理为基础,识别纤维品质改良相关的新基因,研发纤维品质改良转基因棉花,加快我国的棉花纤维品质改良的进程。
3.2 安全性评价和管理策略
转基因棉花的主要环境安全性问题是靶标病虫草的抗性,以及转基因棉花生态系统生物多样性和管理模式的变化对有害生物地位演化的影响。因此,针对转基因抗病虫、耐除草剂棉花等潜在的环境风险问题,我国应加强环境安全性评价及采取相应的风险管理策略。
3.2.1 转基因抗病虫棉花
转基因抗虫棉花安全性评价主要是指转基因抗病虫棉对棉田生态系统靶标生物地位演化、对农业生态系统群落结构和生物多样性的影响以及靶标生物对转基因抗病虫棉花的抗性监测及风险评估[37-38]。因此,要建立转基因抗病虫棉花对天敌种类和数量、经济昆虫的环境风险及其节肢动物群落结构的影响等方面的安全评价体系[39],要明确抗病虫蛋白对棉花病虫害的作用机理和抗性机制[31],发展以分子快速检测为基础的抗性监测平台,针对我国这种特殊的棉花种植模式的特点,对抗性治理因素进行评估[40-42]。针对转基因抗病虫棉花种植的潜在风险,首先要合理地种植转基因抗病虫棉花,延缓靶标生物的抗性进化;其次要积极推广应用基于农业防治和化学防治相结合的非靶标生物的综合防治技术[43-45]。
3.2.2 转基因耐除草剂棉花
杂草抗性和转基因耐除草剂棉花对棉田及周边的植物群落结构和生物多样性的影响是安全性评价的核心内容[46-48]。转基因耐除草剂棉花商业化种植将增加除草剂的使用量,长期使用单一的除草剂必然会导致杂草产生抗性,同时也可能对棉田及周边的植物群落结构和生物多样性产生影响。因此,要建立有效的防控技术和适当的管理措施防止杂草对除草剂产生抗性,通过研究除草剂对植物群落演替的影响,发展以种植转基因耐除草剂棉花为核心的杂草治理技术体系,长期高效地控制杂草的发生与危害[49-51]。
[1] Wu K,Lu Y,Feng H,et al.Suppression of cotton boll wor m in multiple crops in China in areas with Bt toxin-containing cotton[J].Science,2008,321:1676-1678.
[2] Collinge D B,Lund O S,Thordal-Christensen H.What are the pr ospects f or genetically engineered,disease resistant plants?[J].European Jour nal of Plant Pat hology,2008,121:217-231.
[3] Huang J,Pray C,Rozelle S.Enhancing t he cr ops to feed t he poor[J].Nature,2002,418:678-684.
[4] Rajasekaran K,Cary J W,Jaynes J M,et al.Disease resistance conferred by the expression of a gene encoding a synthetic peptide in transgenic cotton(Gossypiu m hirsutu m L.)plants[J].Plant Biotechnology Jour nal,2005,3:545-554.
[5] Lv S,Zhang K,Cao Q,et al.Overexpression of an H+-P Pase gene fr o m Thell ungiella halophil a in cott on enhances salt tolerance and i mproves growth and photosynthetic perf or mance[J].Plant and Cell Physiology,2008,49:1150-1164.
[6] Toenniessen G H,O’Toole J C,De Vries J.Advances in plant biotechnology and its adoption in developing countries[J].Current Opinion in Plant Biology,2003,6:191-198.
[7] Ja mes C.Global stat us of co mmercialized biotech/GM cr ops:2010[M].Brief no.42.ISAAA,It haca,NY.
[8] Bravo A,Likitvivatanavong S,Gill S S,et al.Bacillus thuringiensis:A story of a successf ul bioinsecticide[J].Insect Biochemistry and Molecular Biology,2011,41:423-431.
[9] Sanahuja G,Banakar R,Twy man R M,et al.Bacillus thuringiensis:a cent ur y of research,develop ment and co mmercial applications[J].Plant Biotechnology Jour nal,2011,9:283-300.
[10]Lu Y,Wu K,Jiang Y,et al.Mirid bug outbreaks in multiple crops correlated with wide-scale adoption of Bt cotton in China[J].Science,2010,328:1151-1154.
[11]Lu Y,Qiu F,Feng H,et al.Species composition and seasonal abundance of pestiferous plant bugs(Hemiptera:Miridae)on Bt cotton in China[J].Crop Pr otection,2008,27:465-472.
[12]Wu K,Li W,Feng H,et al.Seasonal abundance of t he mirids,Lygus l ucor u m and Adel phocoris spp.(Hemiptera:Miridae)on Bt cotton in northern China[J].Crop Protection,2002,21:997-1002.
[13]Baum J,Sukuru U R,Penn S R,et al.Identification of hemipteran-active Bacill usthuringiensis cr ystal proteins and evaluation of in pl anta efficacy f or control of Lygus bugs in cotton[C]∥Inter national Congress on Invertebrate Pat hology and Micr obial Control &44t h Annual Meeting of t he Society f or Invertebrate Pat hology,07-11 Aug 2011,Halifax,Nova Scotia,Canada.
[14]Bates S L,Zhao J,Roush R T,et al.Insect resistance manage ment in GM crops:Past,present and f ut ure[J].Nature Biotechnology,2005,23:57-62.
[15]Llewellyn D J,Mares C L,Fitt G P.Field perf or mance and seasonal changes in t he efficacy against Helicover pa ar miger a(Hübner)of transgenic cotton expressing t he insecticidal pr otein vip3 A[J].Agricultural and Forest Ento mology,2007,9:93-101.
[16]Gatehouse J A.Plant resistance towar ds insect her bivores:a dyna mic interaction [J]. New Phytologist,2002,156:145-169.
[17]Mazu mdar-Leight on S,Br oad way R M.Identi fi cation of six chy motrypsin c DNAs fro m larval midguts of Helicover pa zea and Agr otis ipsilon feeding on t he soybean(Kunitz)tr ypsin inhibit or[J].Insect Biochemistry and Molecular Biology,2001,31:633-644.
[18]Wittstock U,Ager bir k N,Stauber E J,et al.Successf ul herbivore attack due to metabolic diversion of a plant chemical defense[J].Proceedings of the National Academy of Sciences of the United States of America,2004,101:4859-4864.
[19]Mao Y,Cai W,Wang J,et al.Silencing a cotton boll wor m P450 monooxygenase gene by plant-mediated RNAi i mpairs larval tolerance of gossypol[J].Nature Biotechnology,2007,35:1307-1313.
[20] Mao Y,Tao X,Xue X,et al.Cotton plants expressing CYP6 AE14 double-stranded RNA show enhanced resistance to boll wor ms[J].Transgenic Research,2011,20:665-673.
[21]Bau m J A,Bogaert T,Clinton W,et al.Control of coleopteran insect pests t hrough RNA interference[J].Nature Biotechnology,2007,25:1322-1326.
[22]Co mai L,Sen L C,Stal ker D M.An altered aro A gene product confers resistance to t he her bicide glyphosate[J].Science,1983,22:370-371.
[23]Nida D L,Kolacz K H,Buehler R E,et al.Glyphosate-tolerant cotton:genetic characterization and pr otein expression[J].Jour nal of Agricultural and Food Chemistry,1996,44:1960-1966.
[24]Tan S,Evans R,Singh B.Her bicidal inhibitors of amino acid biosynthesis and herbicide-tolerant crops[J].Amino Acids,2006,30:195-204.
[25]Keller G,Spatola L,Mccabe D,et al.Transgenic cott on resistant to her bicide bialaphos[J].Transgenic Research,1997,6:385-392.
[26]Blair-Kert h L K,Dotray P A,Keeling J W,et al.Tolerance of transfor med cotton t o gl uf osinate[J].Weed Science,2001,49:375-380.
[27]Wright T R.Biochemical mechanism and molecular basis for ALS-inhibiting her bicide resistance in sugar beet(Beta vul garis)so matic cell selections[J].Weed Science,1998,46:13-23.
[28] Meredit h W R.Cott on yield pr ogress-why has it reached a plateau?[J].Better Crops,2000,84:6-9.
[29]Zhang M,Zheng X,Song S,et al.Spatiotemporal manipulation of auxin biosynt hesis in cott on ovule epider mal cells enhances fiber yield and quality[J].Nat ure Biotechnology,2011,29:453-458.
[30]Wu K,Guo Y.The evol ution of cott on pest management practices in China[J].Annual Review of Ento mology,2005,50:31-52.
[31]Liu C X,Li Y H,Gao Y L,et al.Cotton boll wor m resistance to Bt transgenic cott on:A case analysis[J].Science China Life Science,2010,53:934-941.
[32]Zhang H,Yin W,Zhao J,et al.Early war ning of cotton bollwor m resistance associated with intensive planting of Bt cotton in China[J/OL].PloS ONE,6(8):e22874.
[33]FerréJ,van Rie J.Biochemistr y and genetics of insect resistance t o Bacill us thuringiensis[J].Annual Review of Ento mology,2002,47:501-533.
[34]Zhao J Z,Cao J,Li Y X,et al.Transgenic plants expressing t wo Bacill us t huringiensis toxins delay insect resistance evol ution[J].Nature Biotechnology,2003,21:1493-1497.
[35]Gao Y,Wu K,Gould F,et al.Cr y2 Ab tolerance response of Helicover pa ar miger a (Lepidoptera:Noctuidae)populations fr o m Cr y1 Ac cotton planting region[J].Jour nal of Econo mic Ento mology,2009,102:1217-1223.
[36]An J,Gao Y,Wu K,et al.Vip3 Aa t olerance response of Helicover pa ar migera populations from a Cry1 Ac cotton planting region[J].Jour nal of Econo mic Ento mology,2010,103:2169-2173.
[37]吴孔明.我国Bt棉花商业化的环境影响与风险管理策略[J].农业生物技术学报,2007,15(4):1-4.
[38]Wu K.Monitoring and management strategy f or Helicover pa ar miger a resistance to Bt cotton in China[J].Jour nal of Invertebrate Pat hology,2007,95:220-223.
[39]Yu H,Li Y,Wu K.Risk assess ment and ecological effects of transgenic Bacillus thuringiensis crops on non-target organisms[J].Journal of Integrative Plant Biology,2011,53:520-538.
[40]Gould F.Testing Bt ref uge strategies in the field[J].Nature Biotechnology,2000,18:266-267.
[41]Wu K,Guo Y,Gao S.Eval uation of t he natural ref uge f unction for Helicover pa ar miger a(Lepidoptera:Noctuidae)within Bacill us t huringiensis transgenic cott on gro wing areas in north China[J].Journal of Economic Entomology,2002,95:832-837.
[42]Wu K,Feng H,Guo,Y.Eval uation of maize as a ref uge f or management of resistance to Bt cotton by Helicover pa ar miger a (Hübner)in t he Yellow River cotton-far ming region of China[J].Crop Protection,2004,23:523-530.
[43]陆宴辉,梁革梅,吴孔明.棉盲蝽综合治理的研究进展[J].植物保护,2007,33(6):10-15.
[44]陆宴辉,吴孔明,姜玉英,等.棉花盲蝽的发生趋势与防控对策[J].植物保护,2010,36(2):150-153.
[45]陆宴辉,吴孔明.棉花盲椿象及其防治[M].北京:金盾出版社,2008.
[46]Wolfenbar ger L L,Phifer P R.The ecological risks and benefits of genetically engineered plants[J].Science,290:2088-2093.
[47]Wert h J A,Preston C,Taylor I N,et al.Managing t he risk of glyphosate resistance in Australian glyphosate-resistant cotton pr oduction systems[J].Pest Manage ment Science,2008,64:417-421.
[48]Ramessar K,Peremarti A,Gómez-Galera S.Biosafety and risk assess ment framewor k f or selectable mar ker genes in transgenic crop plants:a case of the science not supporting the politics[J].Trangenic Research,2007,16:261-280.
[49]Valver de B E,Gressel J.A strategy to pr ovide long-ter m control of weedy rice while mitigating herbicide resistance transgene flo w,and its potential use f or ot her crops wit h related weeds[J].Pest Management Science,2009,65,723-731.
[50]Culpepper A S,Yor k A C,Batts R B,et al.Weed management in gluf osinate-and glyphosate-resistant soybean (Gl ycine max)[J].Weed Technology,2000,14:77-88.
[51]Wert h J A,Preston C,Taylor I N,et al.Managing t he risk of glyphosate resistance in Australian glyphosate-resistant cotton pr oduction systems[J].Pest Manage ment Science,2008,64:417-421.
猜你喜欢
今日农业(2022年8期)2022-07-05
少儿科学周刊·儿童版(2022年10期)2022-06-27
今日农业(2022年4期)2022-06-01
山东农业科学(2021年6期)2021-07-16
新高考·英语进阶(高二高三)(2019年1期)2019-06-25
新高考·英语基础(高一)(2019年1期)2019-06-25
中国棉花(2019年8期)2019-01-04
文苑(2018年19期)2018-11-09
浙江农业学报(2017年1期)2017-05-17
上海农业学报(2017年3期)2017-04-10
