
沙棘“生态经济育种”研究Ⅲ——半干旱地区沙棘杂交优势的研究与应用
2011-11-26 06:33:26金争平张吉科温秀凤
水资源开发与管理 2011年3期
金争平,张吉科,温秀凤
(1.水利部沙棘开发管理中心,北京 100038;2.山西大学生命科学学院,太原 030006)
杂交育种增产的主要原因,是因为子一代(F1)新组合中能出现在生长势、生活力、结实性、抗逆性、适应性、产量和品质等方面的表现有超亲现象的单株,表现出明显的杂种优势(heterosis)供我们选择。杂种优势来源于双亲基因的互补、异质等位基因互作、非等位基因互作,加性基因之间、加性与显性基因之间、以及显性基因之间的互作,或这些综合因素累加作用而引起的结果;同时,也是基因群与生境间互作的最终表现。因此,世界上第一次、第二次绿色革命都是依杂交技术获得的优良杂种,大大的提高了作物的单产和品质,减少了人类数量增长引起的粮食饥荒。远缘杂交效果比杂交更为明显和有效,因为杂种所涉及的双亲多基因性状,以及环境差异更大、更多,更易显示出杂种后代的显性、超显性、上位性效应,尤其是上位性效应是杂种优势产量及其构成产量因素的重要遗传基础。[1-3]
在杂种的一生中,优势的表现,是基因 (细胞核基因和细胞质基因)与环境互作过程中遗传力的多组配属性、连续性、时空有序性的表达过程。这个过程体现在营养生长、生殖生长、生理代谢、形态与结构组成、产量形成、品质性状、抗性与生境适应等方面都与亲本有所变异,呈现出复杂的优劣异种多样性[4,5],需要人为地选优与培养,才能达到选优育种的目的。
在前文对沙棘杂交亲本选择研究[6]基础上,本文重点讨论在半干旱地区沙棘杂交F1代杂种优势的表型、生理、品质等方面的研究分析,以便总结经验,提供育种理论与栽培方面相结合的参考依据。
1 材料与方法
实验基地位于内蒙古东胜九成宫水利部沙棘管理中心科研示范基地 (基地概况前文已介绍)。母本沙棘优选了从俄罗斯引种的“太阳”、“楚依”和“优胜”良种,父本是从内蒙古优选的中国沙棘亚种“蛮汗山雄”(它们的性状前文也已介绍)。2000年在坝口通过套袋授粉,获得“太阳×蛮汗山”、“楚依×蛮汗山”和“优胜×蛮汗山”的杂交种子。2001年获得F1代杂种苗,2002年在九成宫基种植,2005年开花结实。在800株 (代号AA)的“太阳杂雌”、300株 (代号AC)的“楚伊杂雌”、200株 (代号AD)的“优胜杂雌”中进行了表型、生理,叶、根结构和部分营养品质的调查与优选研究。
2 结果与分析
2.1 杂种与亲本沙棘营养体表型性状与分析
在特定的生境条件下,沙棘的营养体生长势,主要是由株高、果枝数、新梢数、叶数和叶面积等表型指标所表现。从表1“太阳杂雌”,表2“优胜杂雌”以及亲本营养生长表型特征的8个对比指标看到,杂种沙棘的果枝数、新梢数、新梢叶密度、叶总数、叶面积,都优于双亲,表现出显著的超亲优势 (表型指标大于双亲中的较好值HP)。尤其是新梢数量远超双亲,“太阳杂雌”是母本的3.48倍,是父本的1.18倍;“优胜杂雌”是母本的3.93倍,是父本的1.86倍。新梢和叶数量的增多,为提高结实数量奠定了潜在的基础。新梢多则叶多,单株叶数量,“太阳杂雌”是母本的4.67倍,是父本的1.45倍;“优胜杂雌”是母本的3.28倍,是父本的2.06倍。叶数量和面积的增多,为光合产物的积累奠定了基础。
从表1和表2的三个优株AA54、AD05和AD31的杂种优势来看,他们的果枝数、新梢数、单株叶数和叶面积,超亲优势更为突出,为优良株系的筛选打下了基础。
以上资料表明,这两种杂交后代的基因产生了有利的重组,所表现出的遗传力在营养体的生物学性状上都明显的加大了沙棘个体的体积和生长势,并偏向了父本,也意味着杂种优株偏向了对干旱环境的适应能力,是半干旱地区杂交育种所希望的预期指标。
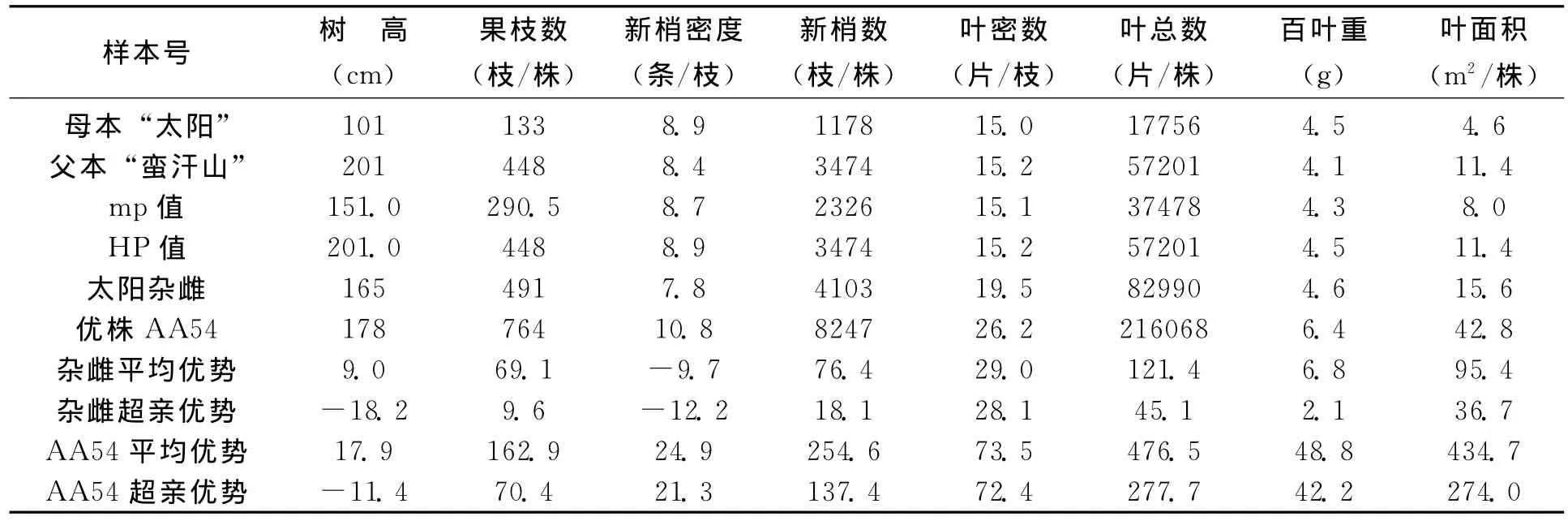
表1 “太阳杂雌”与双亲营养体表型比较及杂种优势 (%)
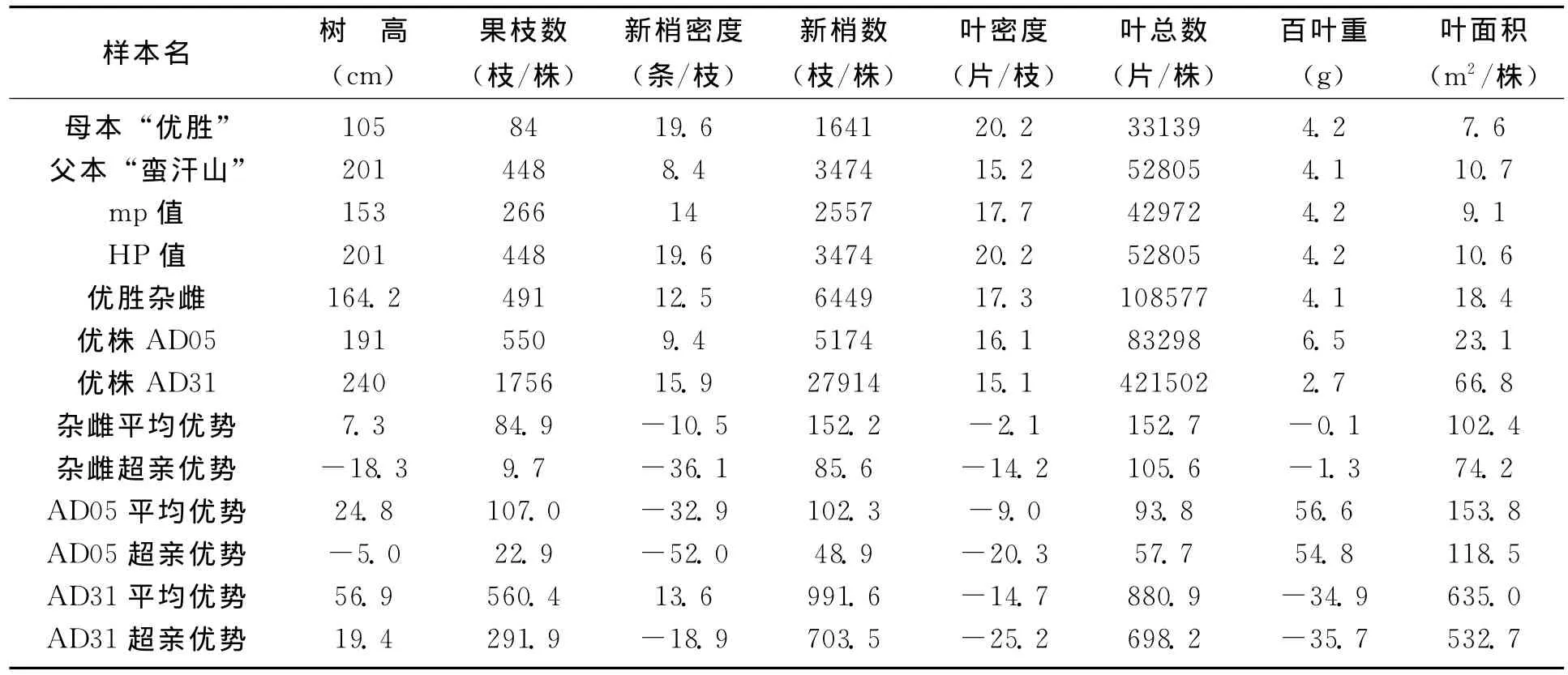
表2 “优胜杂雌”与双亲营养体表型比较及杂种优势 (%)
沙棘植株的营养体中,新梢的生长最活跃,数量最多,新梢来年成为结果枝,结果枝来年成为骨干枝构成树冠。图1反映,株高与新梢数显著正相关,表明株高,冠幅大则新梢多;而图2虽然也反映了株高与结果枝数亦显著正相关,但相关显著性明显低于株高与新梢的关系。这是因为,常年的连续干旱胁迫,导致新梢大量脱落,到新梢停止生长的10月,“太阳杂雌”沙棘平均落梢率高达69.2% (表3),使来年的结果枝数大量减少,又因个体间落梢率的差异,因而影响了株高与结果枝的相关性。可见,干旱限制了营养体的生长发育,造成新梢脱落和结果枝减少,抑制了杂交优势潜能在不利的生境条件下的显性发挥。
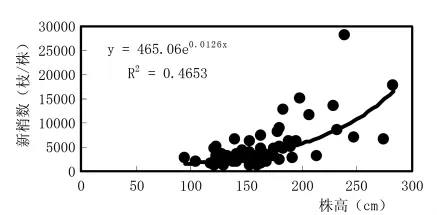
图1 杂雌沙棘株高与新梢数量关系
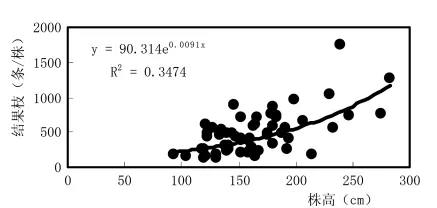
图2 杂雌沙棘株高与结果枝数关系

表3 “太阳杂雌”沙棘6月和10月果枝新梢密度及落梢率 (n=32)
干旱导致新梢脱落连同叶一起脱落,而不脱落的新梢也因干旱影响而落叶,“太阳杂雌”8月下旬的落叶率达15.6% (表4)。

表4 “太阳杂雌”6月和8月果枝叶密度及落叶率 (n=32)
综上所述,杂种沙棘超亲的营养生长势的优势,是父本遗传的显性优势,而干旱环境连续胁迫,显性优势受到了严重的抑制。但遗传潜势仍然保留,因为已形成了新的基因组配,一旦环境有所适应,仍具有增产潜力。因此,在九成宫半干旱地区,沙棘杂交育种是一条显效途径。
2.2 杂种与亲本沙棘生殖体表型性状与分析
从杂雌沙棘和亲本组成生殖体结构表型性状的花序数、果实数、百果重、果产量 (表5、表6)来看,也都显示出不同程度的杂种优势。“太阳杂雌”果枝数的超亲优势达9.7%;果枝数、果总数、百果重和果产量的平均优势分别为69.1%,21.6%,17.5%和123%。“优胜杂雌”果枝数、果实数、百果重、果产量的超亲优势分别达到9.7%,14.1%,6.3%和117.4%。显示了杂交也提高了组成生殖体结构的有利变异。
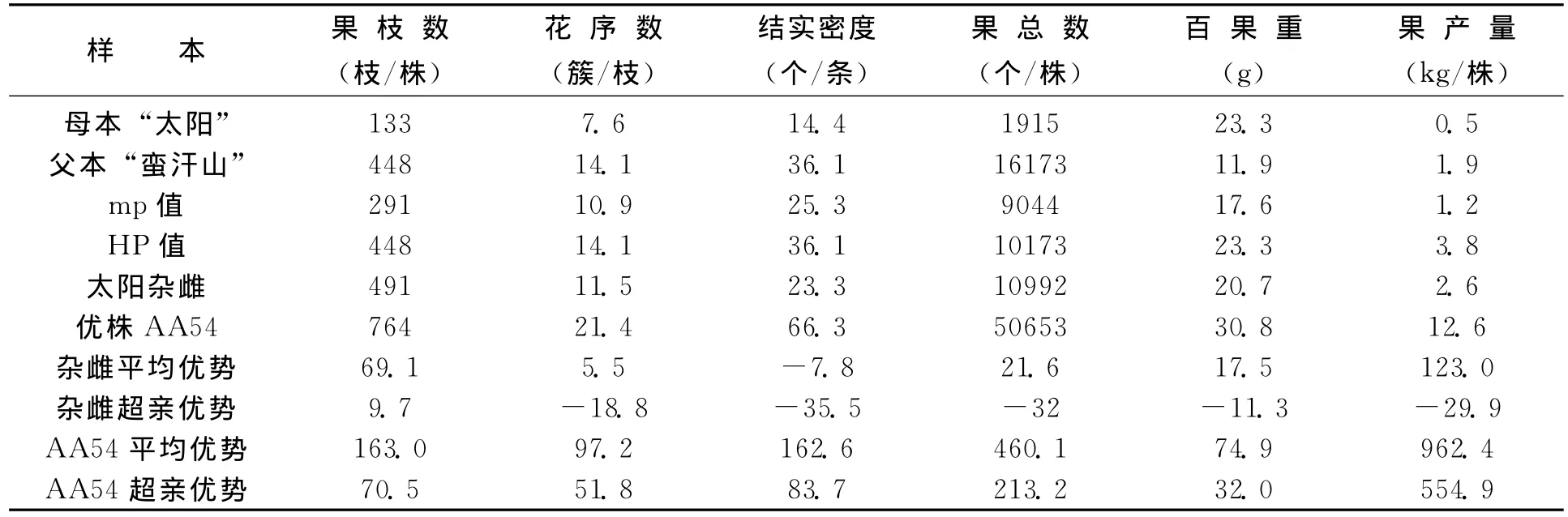
表5 “太阳杂雌”和亲本生殖体表型及杂种优势 (%)
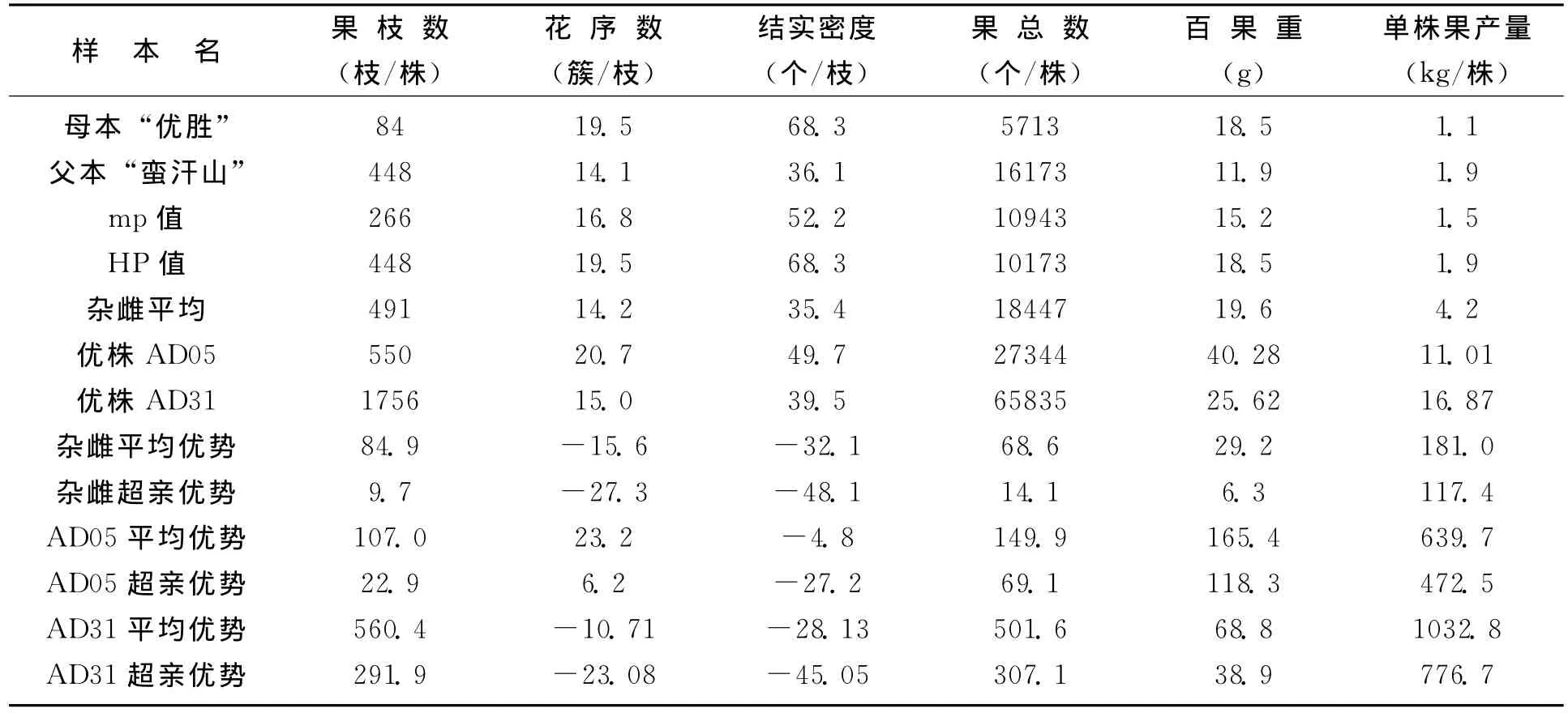
表6 “优胜杂雌”和亲本生殖体表型及杂种优势 (%)
杂交之所以提高果产量的重要原因,主要是杂交显著地增加了新梢数和由新梢长成的果枝数,因而增加了果实数。而花序数、每簇花序果实数、每枝果枝的果实数的杂种优势表现的不显著,是因为它们对干旱更为敏感,果实脱落率更高造成的。调查观测表明,杂雌沙棘生殖生长的潜在杂种优势是显著的,在降水多或者给予较充分灌溉的条件下,落果较轻,果实较大,产量较大地提高。表7反映,“杂雌优54号”在较为湿润且灌溉较多的2008年与最为干旱且灌溉较少的2011年比较,落果率减少了20.4%,而百果重增加了10.79g。
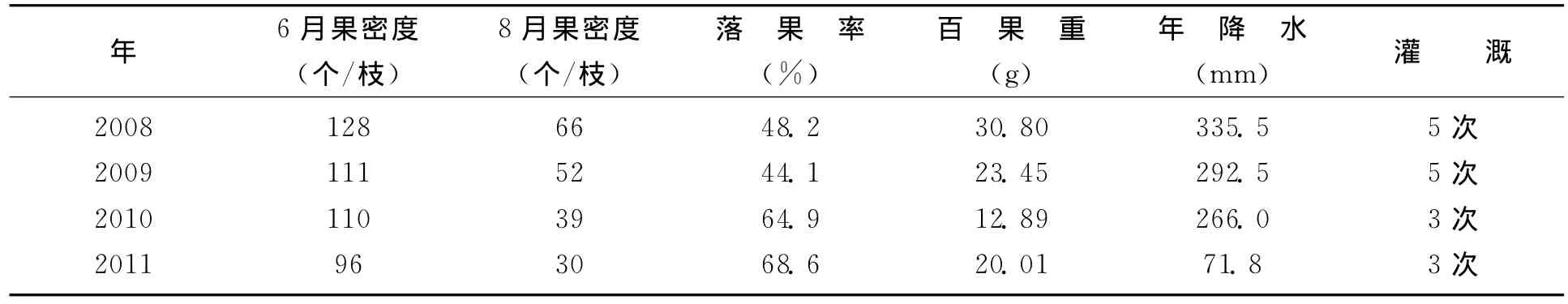
表7 “杂雌优54号”不同水分条件下的落果率和百果重
从影响生殖器官表型基因组配效果来看,“太阳杂雌”优株AA54的表现优秀,因为,它在果枝数、花序数、每簇果实数、每枝果实数、百果重和单株果产量上,都显示出双亲基因的高组配遗传力互补和超显性效应。而“优胜杂雌”优株AD05和AD31,只在果枝数、果实总数、百果重和单株果产量上显示了超亲的显性效应。
2.3 杂种与亲本沙棘生理特性的表现与分析
从3种杂雌沙棘的光合速率日进程 (图3)来看,杂种及父母本最高光合效率都出现在上午的10h,母本最高,父本最低,杂种介于它们之间,比母本光合效率降低了15.8%,比父本光合效率提高了31.5%。表现了杂种优势偏向于母本。更突出的是杂种沙棘没有表现出“光合午休”现象,杂种的光合生理也偏向母本。图4叶气孔导度与光合效率有类似规律。
从图5蒸腾速率日进程看到,在上午10h,蒸腾速率仍然是母本最高,父本最低,杂雌沙棘介于它们之间,但12~16h,杂雌沙棘的蒸腾速率在不断的缓慢上升,这种性状与父本一致,因此,杂雌沙棘的蒸腾特性又表现了其优势偏向于父本,具有抗旱与抗脱水能力。在水分利用效率上 (图6)杂雌沙棘高于母本7.7%,低于父本39.0%,表明杂雌沙棘的水分利用效率的优势偏向于父本,具有节约用水的能力。
总之,杂雌沙棘的光合生理偏向母本,水分利用效率偏向于父本,具有双亲平均遗传优势。这也是从杂交沙棘中选育生态经济型优良单株培育良种所期望的生理特性。
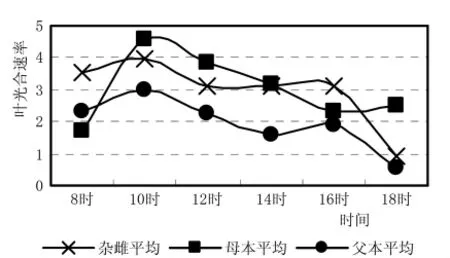
图3 杂雌沙棘和亲本叶光合速率日进程曲线比较
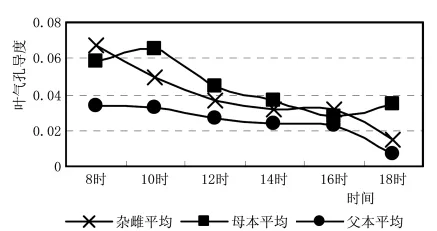
图4 杂雌沙棘和亲本叶气孔导度日进程曲线比较
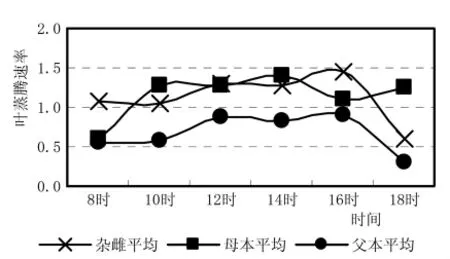
图6 杂雌沙棘和亲本叶蒸腾速率日进程曲线比较
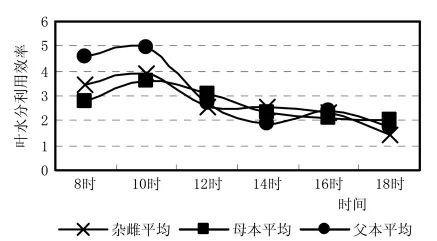
图7 杂雌沙棘和亲本叶水分利用效率日进程曲线
3 结论与讨论
从本文研究的结果来看,在半干旱地区的九成宫基地,沙棘的远缘杂交产生了杂种优势的新组合遗传力,并有部分优良单株供选育,杂种表现出了双亲基因的互补、加性基因间互作,以及综合累加基因群与生境间适应的明显结果。因而,杂种的生长势、生活力、结实性、抗逆性、适应性、产量和品质等方面都具有超亲现象。因此,新杂种沙棘在半干旱地区的九成宫,单产和品质都有了较大幅度的提高和改善。
沙棘杂种优势的主要表现在以下几个方面:
(1)杂交提高了杂种的果枝数、新梢数和叶面积指数,呈现出显著的超亲优势,其生长势偏向了父本遗传,增强了杂种优株对干旱环境的适应能力,为生殖器官的发育与产量奠定了物质和能量的潜在基础。但严重的连续干旱胁迫既导致落梢落叶而影响营养体生长,并影响了生殖体结构花序数、果实数、百果重、单株果产量优势的发挥。尽管如此,杂种还是显示出不同程度的杂种优势,多数表型的平均优势明显,部分表型的超亲优势明显。特别是果实产量的超亲优势显著,比双亲都有大幅度的提高。
(2)杂种的生理表现,其光合强度、气孔导度和蒸腾速率都偏向母本遗传,但水分利用效率偏向于父本遗传。杂种明显地综合了父母本的优良性状,提高了光合强度,提高了水分利用效率,具有双亲平均遗传优势,表现出了杂种增产与节水的效果。更突出的是杂种没有表现出“光合午休”现象,减少了无谓的消耗。
常规杂交育种仍然是当今国内外沙棘界育种的一种重要手段,与其他林木,特别是农作物育种已大力开展与分子育种相结合的方法相比,已经远远落后。希望在今后的沙棘育种中迎头赶上,为沙棘的品种革新做出新的贡献。
[1]刘祖洞 .遗传学 (上册)[M].第2版 .北京:高等教育出版社,1990,286-289.
[2]Xiao JH,Li JM,Yuan LP et a1.1995.Dominance is the major genetic basis of heterosis in flee reveled by Qrn.A nalysis using molecular markers.Genetics.140 (2):745-754
[3]Yu SB,“Jx,Xu CG et a1.1997.Importance of epistasis the genetic basis of heterosis in an elite rice hybrid.Proceedings of the National Academy of Sciences of the USA.94 (17):9226-9231.
[4]吴仲贤 .杂种优势的遗传力理论及其对全球农业的意义[J].遗传学报,2003,30 (3):193-200.
[5]林建丽,朱正歌,高建伟 .植物杂种优势研究进展 [J].华北农学报,2009,24 (增刊):46-56.
[6]金争平,张吉科,温秀凤等 .沙棘“生态经济育种”研究Ⅱ——半干旱地区沙棘杂交亲本比较生态生物学研究 [J].国际沙棘研究与开发,2011,9 (2):23-31.
猜你喜欢
中国饲料(2022年5期)2022-04-26 13:42:38
蔬菜(2021年7期)2021-11-27 03:45:25
东坡赤壁诗词(2020年5期)2020-11-06 07:33:43
小学生时代·综合版(2016年4期)2016-11-19 08:41:24
山西农经(2016年5期)2016-02-28 14:24:37
化工进展(2015年3期)2015-11-11 09:18:44
茶叶通讯(2014年2期)2014-02-27 07:55:39
生物学杂志(2013年1期)2013-04-11 04:19:35
植物营养与肥料学报(2012年1期)2012-10-26 02:49:46
山花(2012年8期)2012-04-29 00:44:03
