
淀山湖浮游藻类增长的氮磷限制性营养研究*
2011-11-08 05:05:28张亚克方焰星张绚璇詹跃武何池全
环境化学 2011年10期
张亚克 梁 霞 方焰星 张绚璇 詹跃武 何池全
(上海大学环境与化学工程学院,上海,200444)
淀山湖浮游藻类增长的氮磷限制性营养研究*
张亚克 梁 霞**方焰星 张绚璇 詹跃武 何池全
(上海大学环境与化学工程学院,上海,200444)
2009年1月至12月对淀山湖浮游藻类生物量及主要营养元素N、P含量进行了监测,并利用藻类增长的生物学评价(NEB)方法对浮游藻类增长的限制性营养元素进行了研究,采用双变量相关性分析了水体中N、P含量与浮游藻类生物量的相关性.结果表明,磷含量与藻类生物量有显著的正相关(Rmax=0.980),添加磷能明显促进藻类生物量的增加,且促进作用随磷浓度的增加而加强;在夏秋之际(7月—9月)蓝藻占优势藻类时,水体中的氮(氨氮)限制了蓝藻的增长,氨氮含量与藻类生物量呈现正相关(Rmax=0.691),硝氮对淀山湖藻类增长促进作用不明显,部分时间表现为抑制作用,磷与硝氮之间存在一定的交互作用.
氨氮,硝氮,磷,浮游藻类,营养限制,淀山湖.
水体富营养化的治理是一个世界性的难题[1-2],在湖泊、水库和缓流态水域中,藻类是主要初级生产者[3-5],氮、磷等营养物在水体中过量累积且外部条件适宜时,容易导致藻类大面积疯长形成水华暴发[6-7].我国大多数湖泊水体中氮磷含量普遍存在超标现象[8-10],是造成水体富营养化的主因.藻类增长的生物学评价(Nutrient Enrichment Bioassay,NEB)是一种有效检测浮游植物营养物限制的主要研究方法,能找出水体中浮游植物增长限制性营养元素[11],并且通过统计试验前后营养盐浓度及藻类叶绿素的变化,可预测以后水体富营养化的程度[12-14],是有效地进行预测、控制藻类增殖的一种方法.
上海市的唯一湖泊淀山湖是上海市郊区饮用水供水水源和水产品养殖基地,其水质具有重要意义[15].但是近年来淀山湖整体水质状况呈逐年下降趋势,淀山湖水体功能定位是Ⅱ类,2001年—2007年淀山湖的水质均未达到Ⅱ类水质标准一般为Ⅳ—Ⅴ类,部分湖区劣于Ⅴ类[16],严重影响到淀山湖生态系统的完整性及上海市郊区饮用水供水安全[17].
本研究通过藻类增长的生物学评价试验确定淀山湖浮游藻类增长的限制性营养元素,为淀山湖富营养化的防治对策及综合治理提供决策依据.
1 材料与方法
1.1 水样采集
于2009年1月至12月每月同一时期在淀山湖四号航标处采集表层水样,采集水样24 h内即进行TN(GB/T 11894—89)、TP(GB/T 11893—89)、叶绿素a[18]等指标的测定,淀山湖采样点位如图1所示.
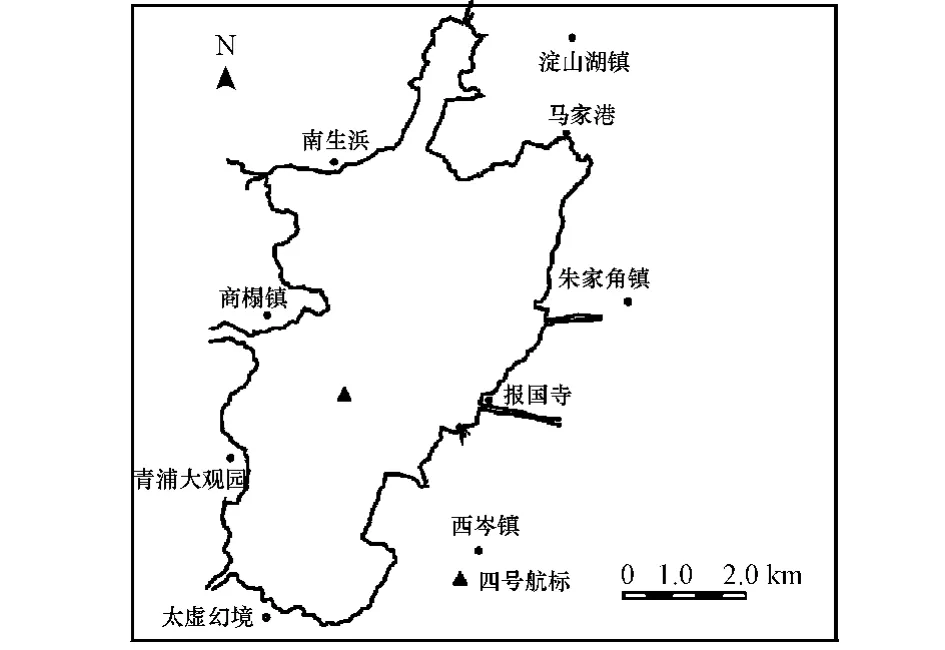
图1 淀山湖地理位置及采样点位置Fig.1 Geagraphic location of Dianshan Lake and sampling sites
1.2 研究方法
根据李比西的最低因子定律(Liebig's Law of the Minimum),植物的生长受它所需要的而存在量最低的营养物质所限制[19-21],藻类增长的生物学评价按照如下营养盐添加方案设计,试验不需对培养水样进行灭菌、过滤和添加藻种等处理,而是直接利用水体中原始的生态群落进行培养[11],可实际的反映含不同时期生物群落的水体在加富营养盐后对浮游植物增长的影响.淀山湖2009年不同月份水体TN、TP含量如表1所示.
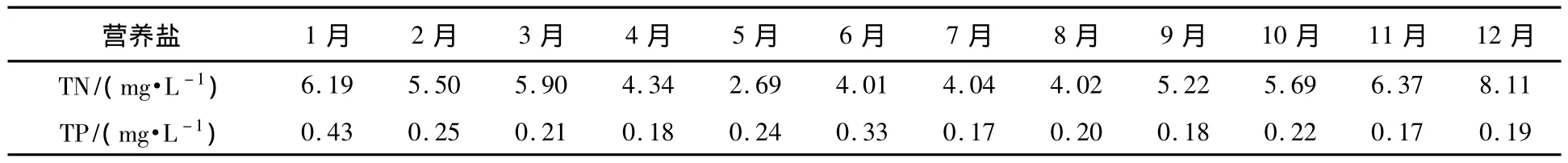
表1 2009年不同月份淀山湖TN、TP含量Table 1 The content of TN and TP in Dianshan Lake in different months
应用标准方法测定淀山湖营养物水平状态,在充分考虑未来淀山湖可能达到的营养物水平的前提下,每个月份试验按原淀山湖水营养物水平(表2)的 0.5倍、1倍、2.0倍添加 P、NO3-N、NH3-N 以及P+NO3-N作为培养液营养物水平,营养盐具体添加方案如表2.量取2 L水样于锥形瓶中按照方案添加不同浓度营养盐在人工气候室内进行培养,每个营养水平设置3个平行样,温度:(25±1)℃;空气湿度70%,光照:4000 Lx,光暗时间比:12∶12,培养一个生长周期.每天取固定量水样用孔径25 mm玻璃纤维滤膜抽滤,后用90%丙酮萃取叶绿素a.萃取液于3500 r·min-1离心20 min,取上清液利用分光光度法测定其叶绿素值.实验方法和计算按规范[22-23]进行.
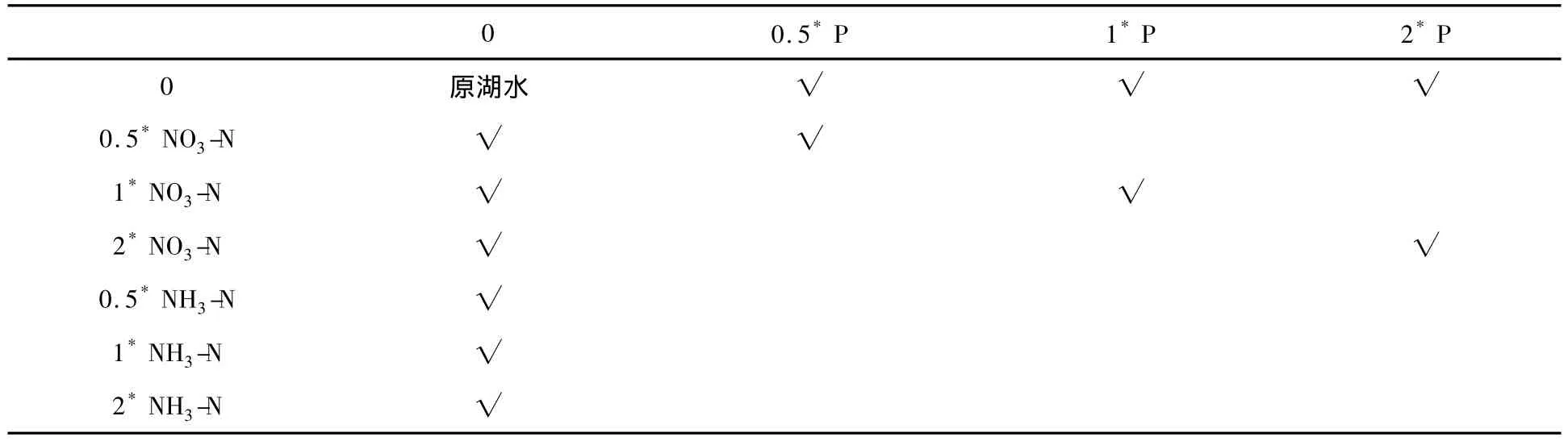
表2 藻类增长实验营养盐添加方案Table 2 Nutrient addition methods of algae growth test
1.3 分析方法
藻类最大现存量是指藻类在一个生长周期内其生物量所能达到的最大值,可用叶绿素a含量X(chl-a)来表征;藻类最大比增长速率是指藻类在一个生长周期内最大的比增长速率.
数据处理为了能够消除不同时期原湖水带来的差异,采用相对比较系数方法来表达添加营养物对藻类增长的影响[24-26].

数据处理均由SPSS 13.0 for Windows软件包完成,对12个月藻类室内培养的增长趋势进行分析,并对不同浓度营养元素与相应藻类叶绿素间的相关性[27]进行分析,以确定不同营养盐添加对藻类生物量的相关性及交互作用的显著性.
2 结果与讨论
2.1 淀山湖叶绿素a、TN、TP的年变化
水体中叶绿素一定程度上可以表征水体中浮游藻类的生物量,是浮游植物现存量的重要指标[28],2009年对淀山湖进行了12个月份的水质监测,其叶绿素a,TN,TP的变化情况如图2所示.
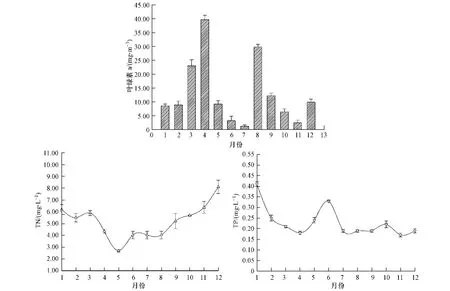
图2 淀山湖叶绿素a,总氮及总磷浓度的月变化Fig.2 The monthly variation of chlorophyll a,TN and TP in Dianshan Lake
王丽卿[29],程曦[30]等研究发现淀山湖历年来在春季和秋季均会出现一个藻类增长高峰期,本实验对淀山湖的监测也得到了类似结果.由图2可以看出2009年淀山湖藻类生物量出现两个高峰,一个出现在春季2—5月,另一个出现在秋季8月份,秋季藻类最高生物量(29.86 mg·m-3)明显低于春季(39.68 mg·m-3).水体中营养物质含量的变化及不同季节藻类优势种群的演替是形成此现象的主要原因.淀山湖藻类的群落演替规律[31]为春季以硅藻和绿藻为主,夏秋季为绿藻和蓝藻,冬季以硅藻为主.由于受到不同季节水温、水文和气象因素的影响,冬季氮磷营养随水体悬浮颗粒物的积累水体中N、P浓度较高[32],但由于气温较低,且冬季光照时间较短,导致冬季浮游藻类并没有出现大量增长;春季(1月至3月)气候转暖,日照时间有所增加且水体中TP、TN均具有较高含量,这给硅藻、绿藻增长提供了有利条件,因此导致在春季4月份时藻类生物量达到峰值;夏季(5月至8月)水体中优势种逐渐转为以蓝藻为主,由于前期藻类大规模增长营养物间的竞争导致消亡氮磷得以释放,水体中氮磷含量有微量上升,特别是氮含量的上升给蓝藻增长提供了有利条件,在8月份藻类的生物量达到另外一个峰值.
2.2 添加营养盐对淀山湖藻类生长的影响
对每月所采水样按照表2方案添加不同浓度氮磷营养盐后进行培养,选择有代表性的几个月份藻类增长情况,其结果如图3.由图3可以看出1月、5月、6月、8月、11月和12月培养实验中,前两天添加不同浓度营养盐组与原湖水组生物量基本相同,从第3天开始添加不同浓度P的藻类生物量开始明显大于原湖水组的藻类生物量,而除8月份外添加不同浓度NH3-N的藻类生物量则低于原湖水组的藻类生物量,表现出明显抑制作用,添加P+NH3-N对藻类增长也表现出明显的促进作用.在淀山湖2月—4月,7月—10月无论添加何种营养盐对藻类增长基本没有影响,生物量一直处于很低水平(叶绿素a<40 mg·m-3).这是由于此时期刚好处于淀山湖藻类大规模增长时期,湖水中氮磷含量趋于饱和,导致添加外源氮磷营养并不能明显促进藻类增长,说明此时期淀山湖浮游植物的增长已基本不再受营养物的限制.而5月—6月、11月—次年1月这几个月份属于夏初和冬初,处于藻类增长的准备时期,添加外源营养元素明显促进了藻类生物量的增加.

图3 藻类增长的生物学评价试验浮游植物增长曲线Fig.3 Phytoplankton growth curves of the nutrient enrichment bioassay test
程曦等[30]通过对浮游藻类群落的长期连续监测数据进行分析后发现淀山湖浮游藻类存在明显的群落演替,主要表现为春季硅藻、绿藻,夏秋季蓝藻以及冬季硅藻为主的特点.由于不同种类藻类细胞的元素组成存在着差异,且不同种藻类对营养盐的生理响应不同,因此藻类群落演替也是导致本实验不同月份藻类增长趋势出现显著差异的一个重要原因.特别是试验中发现8月份添加氮磷都增加了藻类生物量,8月份淀山湖藻类以蓝藻为主且蓝藻自身具有固氮作用,因此导致8月份添加氮磷对藻类增长都具有明显的促进作用.
2.3 添加不同 NH3-N、NO3-N、P下藻类增长的的氮磷限制分析
为了能够消除不同时期原湖水带来的差异,采用相对比较系数方法来表示添加不同营养元素藻类生物量与空白间的差值,其中原湖水的结果均表达为0%,结果如图4所示.
由图4可以看出,全年除了4月、6月份外其余10个月份添加P后藻类相对比较系数大部分在40%与105%之间,添加P后浮游植物生物量远大于原湖水.在2月、3月和8月添加NH3-N和NO3-N后藻类生物量高于原湖水,其余月份添加NH3-N和NO3-N则并没有表现出对藻类增长的促进作用,相反部分时间添加NH3-N和NO3-N抑制了藻类的增长,如4月、5月和6月3个月份.全年除了4月、5月和6月添加NO3-N+P对藻类增长具有抑制作用外,其余月份添加NO3-N+P对藻类增长都体现出不同程度的促进作用,表明在全年中共同添加NO3-N和P时部分时间表现出对浮游藻类增长的促进作用,部分时间则表现出抑制作用,二者之间存在一定的交互作用.

图4 添加不同营养盐下藻类增长相关系数的变化Fig.4 The variation of correlation coefficient in different nutrient addition methods
考虑N、P添加量对藻类增长的影响,可以看出在4月、5月和6月藻类大规模增长时不管添加何种营养物对藻类增长均是抑制作用;其余月份除了1月外,添加P明显的促进了藻类的增长,而且是随着添加量的增大这种促进作用越明显,表明P是淀山湖藻类增长的限制因子;在2月、3月份表现出添加N能促进藻类增长,7月、8月和9月则表现出添加NH3-N能明显促进藻类生物量的增加,添加不同量NO3-N并没有明显促进藻类的增长,相反还出现不同程度的抑制作用;添加NO3-N+P对藻类增长也表现出促进作用,如2、3月表现为大于单一添加P营养物的促进作用,其余月份则表现为缓和了NO3-N,NH3-N营养物对藻类增长的抑制作用,这从另一方面说明了P对淀山湖藻类增长的促进作用.
2.4 营养盐对藻类生长平均最大现存量和平均最大特定增长率的影响
添加不同种类和不同水平营养盐试验组藻类平均最大现存量(Xmax)和平均最大特定增长率(Lmax)见表3.从表3可以看出藻类现存量全年中除4月、9月、10月添加P及NO3-N+P增加不明显外,其余月份添加P及NO3-N+P藻类的现存量明显大于原湖水组,最大藻类现存量出现在8月份,单独添加P组为144.92 mg·m-3,添加 NO3-N+P 组为 139.05 mg·m-3;全年中添加 NO3-N 及 NH3-N 营养组除 3 月,8 月藻类的现存量略大于原湖水组,其余月份始终没有较高的现存量均低于原湖水组,最大藻类现存量也出现在8 月份,单独添加 NO3-N 组为 110.53 mg·m-3,添加 NH3-N 组为 110.63 mg·m-3;说明 N 的加入没有明显促进淀山湖藻类的繁殖增长,而在原湖水中所添加的P营养组,最终都有较大的现存量,因此与N相比,P的加入明显促进了淀山湖藻类繁殖增长.
全年中添加P及NO3-N+P组的藻类平均最大增长率变化与藻类现存量变化类似,除4月、9月、10月藻类增长率增加不明显外,其余月份藻类的增长率明显大于原湖水组;在2月、3月、8月、11月添加NO3-N及NH3-N营养组藻类的最大增长率也大于原湖水组;说明对P和N对淀山湖藻类的生长速度都有促进作用,但P对藻类最大增长率的影响显著,说明P是淀山湖藻类生长繁殖的限制因子;同时由于4月、9月、10月这3个月份淀山湖藻类处于大规模增长的后期,湖水中各营养盐处于饱和状态,藻类普遍趋于衰亡,因此在4月、9月、10月添加各营养盐藻类增长出现了负增长率.
2.5 添加不同 NH3-N、NO3-N、P下对藻类增长的的相关性分析
由数据分析可知,在淀山湖对藻类生长起着限制作用的因子可能是磷,进一步对添加P、NH3-N、NO3-N下的藻类叶绿素含量进行相关分析如表4所示.
由表4知,大部分添加不同浓度P与藻类叶绿素之间在0.05的水平上差异显著,添加P对于2月、3月、7月—9月和11月、12月份水样的藻类叶绿素含量呈显著性正相关,说明P是淀山湖藻类增长的第一限制性元素,添加P能明显促进淀山湖的藻类增长;添加NH3-N对于4月份和5月份水样中藻类叶绿素含量呈显著性负相关,在7月—9月份呈现显著性正相关;添加NO3-N对于5月、6月、9月、12月份的水样中藻类叶绿素含量也呈显著性负相关,说明N不是淀山湖藻类增长的限制性元素,相反增加NO3-N含量表现出对藻类增长具有一定的抑制性作用.


表4 淀山湖全年藻类叶绿素含量相关性分析Table 4 The correlation analysis of full year algae chlorophyll in Dianshan Lake
3 结论
藻类增长生物学评价实验表明,磷是淀山湖浮游植物增长的第一限制性因子,添加磷能明显促进藻类生物量的增加,且促进作用随磷浓度的增加而加强;在夏秋之际蓝藻占优势藻类时,水体中的氮(氨氮)限制了蓝藻的增长,硝氮对淀山湖藻类增长促进作用不明显,部分时间表现为抑制作用,磷与硝氮之间存在一定的交互作用,同时添加磷和硝氮对藻类增长有一定的促进作用.
[1]Shanna Chen,Xiaolan Chen,Yan Peng,et al.A mathematical model of the effect of nitrogen and phosphorus on the growth of blue-green algae population[J].Applied Mathematical Modeling,2009,33:1097-1106
[2]Schindler D W,Heckey R E,et al.Eutrophication:More Nitrogen Data Needed[J].Science,2009,324:721-723
[3]刘信安,湛敏,马艳娥.三峡库区流域藻类生长与营养盐吸收关系[J].环境科学,2005(4):96-99
[4]Walter Mulbry,Shannon Kondrad,Carolina Pizarro,et al.Treatment of dairy manure effluent using freshwater algae:Algal productivity and recovery of manure nutrients using pilot-scale algal turf scrubbers[J].Bioresource Technology,2008(99):8137-8142
[5]Soyupak S.,Mukhallalati L,Yemisen D,et al.Evaluation of eutrophication control strategies for the Keban Dam reservoir[J].Ecological Modeling,1997(97):99-110
[6]李辉,潘学军,史丽琼,等.湖泊内源氮磷污染分析方法及特征研究进展[J].环境化学,2011,30(1):281-285
[7]Lars Håkanson,A general process-based mass-balance model for phosphorus eutrophication as a tool to estimate historical reference values for key bioindicators,as exemplified using data for the Gulf of Riga[J].Ecological Modeling,2009(220):226-244
[8]李建平,吴立波,戴永康,等.不同氮磷比对淡水藻类生长的影响及水环境因子的变化[J].生态环境,2007,16(2):342-346
[9]李姗姗,郭沛涌,吴龙永,等.厦门市石兜坂头水库水体生物有效磷与叶绿素a分布特征及其相关性[J].环境化学,2011,30(2):483-486
[10]Ao Jingqiao,Chen Qiuwen,Chen Yongcan.Three-dimensional eutrophication model and application to Taihu Lake,China[J].Journal of Environmental Sciences,2008,20:278-284
[11]吴雪峰,程曦,李小平.淀山湖浮游植物营养限制因子的研究[J].长江流域资源与环境,2010,19(3):292-298
[12]李桂欣.AGP试验在珠江口监测的应用[D].广州:中国海洋大学,2006.6-11
[13]李坤阳,储昭升,金相灿,等.巢湖水体藻类生长潜力研究[J].农业环境科学学报,2009,28(10):2124-2131
[14]William M,Lewis J R,Wayne A,et al.Control of Lacustrine Phytoplankton by Nutrients:Erosion of the Phosphorus Paradigm[J].Internat Rev Hydrobiol,2008,93:446-465
[15]施玮,吴和岩,赵耐青,等.淀山湖水质富营养化和微囊藻毒素污染水平[J].环境科学,2005,26(5):55-61
[16]郑晓红,汪琴.淀山湖水质状况及富营养化评价[J].环境监测管理与技术,2009,21(2):69-70
[17]张鼎国,杨再福.淀山湖生态环境的演变与对策[J].水利渔业,2006,26(1):61-63
[18]国家环境保护总局水和废水监测分析方法编委会.水和废水监测方法(第4版)[M].北京:中国环境科学出版社,2002
[19]Legovic T,Cruzado A.A model of phytoplankton growth on multiple nutrients based on the Michaelis-Menten-Monod uptake,Droop's growth and Liebig's law [J].Ecological Modeling,1997,99:19-31
[20]Hugo Antonius,Van den,Berg.Multiple nutrient limitations in unicellular:reconstructing Liebig's law [J].Mathematical Biosciences,1998,149:1-22
[21]尹澄清,兰智文.富营养化水体中藻类生长限制因素的确定及其应用[J].环境化学,1993,12(5):380-384
[22]国家环保局《水生生物监测手册》编委会编.水生生物监测手册[M].南京:东南大学出版社,1993
[23]American Public Health Association.Standard Methods for the Examination of Water and Wastewater[M].15th Ed.New York.1980
[24]Arar E J,Collins G B.In Vitro determination of chlorophyll a and pheophytin a in marine and freshwater algae by fluorescence(Method 445.0) [R].National Exposure Research Laboratory,Office of Research and Development,U.S.Environmental Protection Agency,1997
[25]Goldman C R,Wetzel R G.A study of t he primary productivity of Clear Lake,Lake County[J].California Ecology,1963,44(2):283-294
[26]Orne A J,Goldman C R.Nitrogen fixation in Clear Lake,California.1.Seasonal variation and the role of the heterocyst[J].Limnol Oceanogr,1972,17(5):678-692
[27]李志辉,罗平.SPSS for Windows统计分析教程(第二版)[M].北京:电子工业出版社,2003
[28]吴洁,钱天鸣,虞左明.西湖叶绿素a周年动态变化及藻类增长潜力试验[J].湖泊科学,2001.13(2):143-148
[29]王丽卿,张军毅,王旭晨,等.淀山湖水体叶绿素a与水质因子的多元分析[J].上海水产大学学报,2008,17(1):59-64
[30]程曦,李小平.淀山湖氮磷营养物20年变化及其藻类增长响应[J].湖泊科学,2008,20(4):409-419
[31]由文辉.淀山湖着生藻类群落结构与数量特征[J].环境科学,1999,20:59-62
[32]林建伟,朱志良,赵建夫,等.负载硝酸盐有机改性沸石抑制底泥氮磷释放的效果及机制研究[J].环境科学,2008,29(2):356-361
STUDY ON THE LIMITING NUTRIENT ELEMENT OF PHYTOPLANKTON GROWTH IN DIANSHAN LAKE
ZHANG Yake LIANG Xia FANG Yanxing ZHANG Xunxuan ZHAN Yuewu HE Chiquan
(School of Environmental and Chemical Engineering,Shanghai University,Shanghai,200044,China)
The biomass of phytoplankton and content of N,P in Dianshan Lake were investigated from January,2009 to December,2009.The limiting nutrient element of phytoplankton growth was analyzed by nutrient enrichment bioassay experiment,and the correlation between biomass of phytoplankton and content of N,P was investigated by bivariate correlation analysis.The results showed that the content of phosphorus was positively correlated with the biomass of phytoplankton(Rmax=0.980).Addition of phosphorus obviously boosted biomass of phytoplankton,and the facilitation was stronger with large addition of phosphorus.The growth of cyanobacteria was restricted by ammonia nitrogen in water in the summer and fall,when cyanophyta was in predominance.The content of ammonia nitrogen was positively correlated with the biomass of phytoplankton(Rmax=0.691).There was no significant enhancement on the biomass of phytoplankton by the nitrate in Dianshan lake,which sometimes caused the inhibition of phytoplankton.There were effects of interactions between nitrate and phosphorus.
ammonia nitrogen,nitrate,phosphorus,phytoplankton,nutrient limitation,Dianshan Lake.
2010年11月23日收稿.
* 上海大学创新基金项目(No.A.10-0111-08-006)资助.
**通讯联系人,E-mail:asituva@shu.edu.cn
猜你喜欢
环境工程技术学报(2021年1期)2021-01-07 17:14:28
中国甜菜糖业(2018年4期)2018-12-25 07:20:30
水利技术监督(2017年6期)2017-12-19 13:28:17
中国诗歌(2016年6期)2016-11-25 23:54:04
绿色科技(2016年16期)2016-10-11 06:44:40
广东海洋大学学报(2015年3期)2015-12-22 10:05:28
应用海洋学学报(2015年3期)2015-11-22 07:39:10
城市建设理论研究(2014年11期)2014-04-21 00:36:00
华东师范大学学报(自然科学版)(2014年6期)2014-02-27 13:40:57
湖泊科学(2014年2期)2014-02-27 10:29:43
