
不同整地方式对小兴安岭低质林生物多样性的影响
2011-09-06 01:14:26吕海龙董希斌
森林工程 2011年6期
吕海龙,董希斌,2
(1.东北林业大学工程技术学院,哈尔滨 150040;2.黑龙江省森林持续经营与微生物工程重点实验室东北林业大学,哈尔滨 150040)
整地是改善立地条件,提高造林成活率,加快林木生长的重要措施,整地方式多种多样,不同的整地方式,林木的生长情况各不相同[1]。合理的整地不仅可以改善土壤的理化性质,而且是接纳雨雪、拦截径流、蓄水保墒、保持水土的重要措施。特别是在干旱地区,细致征地对提高造林成活率,促进幼林的生长发育具有十分重要的意义。无论是哪种整地方式,都会对森林环境产生不同程度的扰动,环境扰动的存在,通过对不同物种种群的影响,进而改变了群落生境,而生境的差异又可以作用到物种间的竞争和共存模式[2],进而导致了植物群落物种多样性的显著差异[3]。在任何生态类型中,植物种的丰富度是决定物种多样性的主要因子,分析植物群落物种丰富度是物种多样性研究的基础[4]。森林生物多样性是森林生态系统稳定性的重要体现,是森林生态系统演替的外部反映,是森林生态系统功能的衡量标准[5],不同森林类型的林下植物群落在物种组成上具有较大差异,这种差异可以通过林下植物多样性指标来反映,植物多样性指标一般包括物种丰富度指数、多样性指数和均匀度指数等,其中不同指数从不同角度反映了物种多样性的变化,而各种指数之和可以反映物种多样性的总体变化[6]。在多次过伐及自然灾害的多方面影响下,小兴安岭林区形成大面积郁闭度较低,林相衰败的残次林,造成该地区的林分经济和生态效益降低[7-8]。本研究以小兴安岭林区的低质林为研究对象,在带岭林业局和铁力林业局的低质林设置试验样地,对低质林试验样地采用不同的整地方式,从生物多样性的角度探讨低质林区合理的整地方式,为低质林分的改造和经营以及森林生物多样性恢复的研究提供新的依据及参考。
1 研究区自然概况
在小兴安岭林区内选取典型的低质林4个试验区 其中 试验区Ⅰ 试验区Ⅱ和试验区Ⅲ设置在带岭林业局——红光林场。试验区Ⅰ设置在红光林场四线沟的300林班内,公里坐标 (0495068,5235642),是以冷杉为主的针阔混交林,树种组成为4冷3云3枫,平均林龄35a,平均树高14 m,平均坡度10°,林内灌丛覆盖度为60%,主要有榛柴和忍冬等;林下主要指示性植物有苔草、蕨类等,多度为70%;土壤为暗棕壤,厚度30cm。试验区Ⅱ设置在红光林场四线沟的301林班内,公里坐标 (0494766,5234811),是以冷杉为主的针阔混交林,树种组成为4冷3云3枫,平均林龄25a,平均树高12 m,平均坡度5°,林内灌丛覆盖度为85%,主要有榛柴、忍冬等;林下主要指示性植物有苔草、蕨类等,多度为20%;土壤为暗棕壤,厚度30cm。试验区Ⅲ设置在红光林场二线沟的285林班内,公里坐标 (0493912,5230377),是以云杉为主的针阔混交林,树种组成为4云3冷3枫,平均林龄35a,平均树高14 m,平均坡度10°,林内灌丛覆盖度为60%,主要有榛柴、忍冬等;林下主要指示性植物有苔草、蕨类等,多度为
土壤为暗棕壤 厚度 试验区Ⅳ设置在铁力林业局——卫东林场的187林班内,公里坐标 (0445321,5248633),是以桦树为主的阔叶混交林,树种组成为3白3枫2水1椴1色,平均林龄30a,平均树高15 m,平均坡度5°,林内灌丛覆盖度为40%,主要有榛柴等;林下主要指示性植物有苔草等,多度为40%;土壤为暗棕壤,厚度50cm。
2 研究方法
2.1 试验区设计方法
试验区实验样地郁闭度均为0.4,且均采用林下造林、人工更新、抚育年限为五年 (抚育次数为2.2.1.1.1);试验区Ⅳ中的营造红松云杉混交林的混交比例为3∶1,其他各设计方法见表1。在试验区Ⅰ、试验区Ⅱ、试验区Ⅲ和试验区Ⅳ每个试验区相邻未改造的低质林内,分别取保留样地D1、D2、D3和D4并调查其中的生物多样性数据,与相应的试验区改造样地作对比,分析改造后样地的生物多样性与原低质林保留样地的生物多样性之间的差异。

表1 试验区设计表Tab.1 Design table of test area
2.2 多样性指数的计算
传统的生物多样性测度指标有:Simpson指数、Shannon-wiener指数、Pielou均匀度指数、Margalef指数、Gleason指数等[9]。均匀度是指样地中各个种的多度的均匀程度,即每个种个体数间的差异,其计算常用观察多样性和最高多样性的比值来表示,最高多样性即所有种的多度相等时的多样性,据此可导出均匀度的计算式[10-16]。

式中:pi=niN,代表第i个物种的相对多度。
(2)本文采用Pielou均匀度指数J:

式中:S 为样方中物种数[17-18]。
2.2.1 乔木层物种多样性
依据调查数据运用公式 (1)、(2)计算得到各灌木层生物多样性指数见表2。

表2 乔木层生物多样性指数Tab.2 Biodiversity of tree layer
在造林更新的改造方式下,对试验区内的低质林林木进行清理后试验区改造样地内主要树种是红松、枫桦、云杉、冷杉、水曲柳、和紫椴等,还有少量的五角槭、黄菠萝、青楷槭和白桦等。由表2可见 试验区Ⅱ样地的乔木层物种多样性指数(2.05)和均匀度指数 (0.91)总体最高,已超过原始阔叶红松林的乔木层物种多样性指数 (1.89)和均匀度指数 (0.82),这是因为,该改造样地内林木的种类较多,且林木数量少分布零散却均匀,但实验区内出材率较高的林木很少,远远低于原始阔叶红松林。每个试验改造样地的乔木层生物多样性指数与其保留样地总体对比可见,不同整地方式下改造样地的乔木层物种多样性指数总体高于试验区的保留样地,且每个改造样地的乔木层均匀度指数比与其对应的保留样地的乔木层均匀度指数都高;改造后的样地的乔木层物种多样性指数和均匀度指数都接近或高于原始阔叶红松林。说明在改造样地的过程中,人为保留林木的种类、数量及分布位置都非常合理,对乔木层生物多样性恢复有促进作用。但是出材率高的林木很少,故在选择性的保留林木的基础上补植幼苗是有必要的,既降低了对森林环境的扰动,又促进了人工更新。
2.2.2 灌木层物种多样性
依据调查数据运用公式 (1)、(2)计算得到各灌木层生物多样性指数见表3。
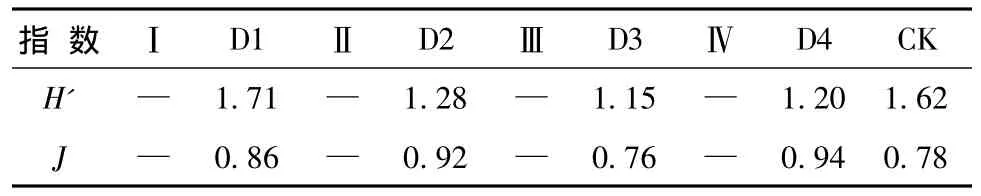
表3 灌木层生物多样性指数Tab.3 Biodiversity of shrub layer
由于试验区样地是2011年改造的,更新带中的灌木都被清理,所以改造样地中的灌木层生物多样性没有调查数据。试验区保留样地和原始阔叶红松林的灌木层植物主要有榛子、暴马丁香、忍冬、珍珠梅、刺五加、三颗针、悬钩子等,还有少量的接骨木、茶藨子、溲疏、醋粟、刺老芽和瘤枝卫矛等。由表3可见,试验区Ⅰ的保留样地D1样地灌木层的物种多样性指数 (1.71)和均匀度指数 (0.86)均最高,且高于原始阔叶红松林,这是由于,D1样地内乔木层林木稀少,灌木层植物缺少强大的乔木层植物的生存竞争,使得保留样地内的通风、光照、温度、土壤湿度等因素都适合灌木层植物生长,导致D1样地的灌木层物种多样性指数过高。试验区的保留样地与原始阔叶红松林总体对比得,保留样地的灌木层物种多样性指数普遍低于原始阔叶红松林,而灌木层均匀度指数普遍高于原始阔叶红松林,这说明保留样地内乔木层林木稀少,使得灌木层植物分布均匀 而保留样地内灌木层植物的种类远远低于原始阔叶红松林,因此反映了试验区内低质林的生态环境与原始阔叶红松林有较大的差距,低质林生态系统恢复任务十分艰巨,必须及时并合理的对低质林进行改造,促进低质林生物多样性的恢复。2.2.3 草本层物种多样性
依据调查数据运用公式 (1)、(2)计算得到各草本层生物多样性指数见表4。

表4 草本层生物多样性指数Tab.4 Biodiversity of herb layer
试验区内草本层植物主要有毛缘苔草、羊胡苔草、蕨、小叶樟、蚊子草、山苞米、三楞草、毛茛、猪殃殃、荨麻、侧金盏花、蒿、延尾凤毛菊、北重楼、唐松草、野豌豆、山尖子和木贼等,其中试验区内部分区域还有独活、延胡索、单叶舞鹤草、朝鲜顶冰花、河清花、菟葵、早熟禾、大叶柴胡、驴蹄菜、耳叶蟹甲、萎陵菜、附地菜、香茶菜、落新妇、金腰子、堇菜、蓍草、草乌、天南星、五福草、假升麻和花忍等。由表4可知,D4样地的草本层物种多样性指数和均匀度指数均高于原始阔叶红松林,且试验区内每个样地的草本层物种多样性指数都很高,这也反映了在低质林实验区内的阳光、温度、通风、温度、土壤湿度等自然因素都有益于草本植物生长,使得草本植物生长茂盛且种类增多;按整地方式将实验区内样地分为揭草皮子整地方式一组和暗穴整地方式一组,对这两种整地方式进行比较,将两组整地方式的各个实验区内的改造样地与其对应的保留样地的草本层物种多样性指数H'和均匀度指数J分别取比值的平均值得,揭草皮子整地方式的草本层物种多样性指数H'比值的平均值为0.984和均匀度指数J比值的平均值为0.971,暗穴整地方式的草本层物种多样性指数H'比值的平均值为0.915和均匀度指数J比值的平均值为0.934。草本层生物多样性指数比值的平均值越接近于1,反映这种整地方式对草本层生物多样性的负面影响越小,揭草皮子整地方式改造样地与其保留样地草本层生物多样性指数比较接近于1,即揭草皮子的整地方式方法比暗穴整地方式方法对草本层生物多样性的负面影响小。
根据表2、表3、表4的生物多样性数据,对乔木层 灌木层和草本层及乔木层和灌木层的物种多样性指数H'值分别作相对比较 (如图1所示):在乔木层、灌木层、草本层的物种多样性指数H'值的相对比较中 (如图1左侧所示),保留样地D1与原始阔叶红松林最为接近,说明初始条件下该低质林乔木层、灌木层、草本层物种多样性的匹配比较合理;就乔木层和草本层的物种多样性指数H'值的相对比较中 (如图1所示),改造样地内的Ⅰ样地与原始阔叶红松林最为接近,揭草皮子整地方式下的改造样地 (Ⅰ、Ⅱ和Ⅲ)比暗穴整地方式下的改造样地 (Ⅳ)的乔木层和草本层的物种多样性指数H'值的比例更接近原始阔叶红松林,且揭草皮子整地方式改造样地与保留样地的乔木层和草本层的物种多样性指数H'值的比例比较接近,说明揭草皮子整地方式优于暗穴整地方式,在低质林改造整地方式的选择中,揭草皮子整地方式和暗穴整地方式相比,可优先考虑选择揭草皮子整地方式。

图1 乔木层、灌木层、草本层H'的相对值Fig.1 Relative value of H'in trees layer,shrub layer and herb layer
2.3 各实验区物种多样性恢复主成分分析
为了从整体分析各个试验区生物多样性的初始效果,以各试验区内改造样地和保留样地的物种多样性指数和均匀度指数为基础,运用主成分分析(Principalcomponent analysis),计算各试验区的综合得分。主成分特征值见表5。
由表5可知,前两个公因子特征值均大于1,且第1主成分贡献率为66.077%。前两个主成分累计解释率达83.302%,因此能够充分描述各试验区生物多样性的初始情况。
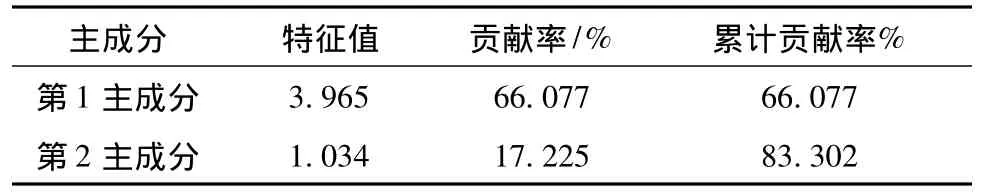
表5 特征值解释Tab.5 Eigenvalue explains
由表 可以看出草本层 和 在第一公因子(F1)上有很大载荷,定义F1为草本层多样性指数因子,灌木层和乔木层多样性指数在第二公因子(F2)上有很大载荷,F2定义为灌木层和乔木层多样性指数因子,两个公因子分别从不同方面反映了各试验区的生物多样性初始情况,单独一个公因子不能反映某一调查区的初始情况,按各公因子对应的贡献率为权数计算:

式中:F为综合得分;λ1为第一主成分贡献率;λ2为第二主成分贡献率;S1为第一主成分因子得分;S2为第二主成分因子得分。
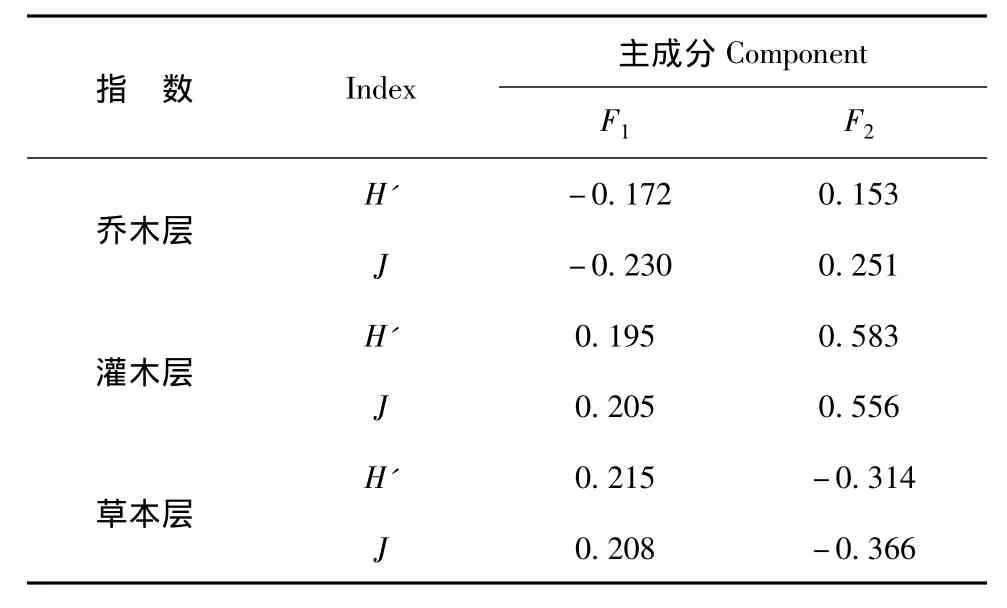
表6 因子载荷表Tab.6 Factor loading table
按照公式 (3),依据各因子得分计算各试验区多样性初始情况综合得分,见表7。

表7 各试验区综合得分Tab.7 Comprehensive scores of each test area
由表7可见:红光林场301林班的保留样地(D2)的综合得分 (0.52)最高,红光林场300林班的保留样地 (D1)的综合得分 (0.49)次之;原始阔叶红松林 的综合得分 最高;而改造带的综合得分均为负值,说明试验区改造样地距离恢复达到原始阔叶红松林还需要较长的时间。在揭草皮子整地方式改造样地的综合得分均比暗穴整地方式改造样地高,揭草皮子整地方式比暗穴整地方式对森林生境扰动小。
3 结论与讨论
本文对小兴安岭低质林试验样地的生物多样性进行调查,应用主成分分析的方法,以原始阔叶红松林作对照,对不同整地方式下的低质林生物多样性综合对比发分析的研究中发现:在乔木层中,为了使改造后的森林生物多样性尽快恢复,选择性的保留乔木层出材率高的林木,有利于限制灌木层植物和草本层植物的过盛生长,从而降低改造林地内灌木层和草本层植物对补植幼苗的影响,有益于低质林乔木层生物多样性的快速恢复,在改造样地的过程中,人为保留林木的种类、数量及分布位置都非常合理,对乔木层生物多样性恢复有促进作用;在灌木层中,由于多年的开发利用及自然灾害的破坏,使得乔木层林木稀少,导致灌木层植物分布均匀,但由于生存环境被破坏,因此灌木层植物的种类比原始阔叶红松林少;在草本层中,从草本层生物多样指数总体来看,改造样地的草本层生物多样指数普遍低于保留样地,说明低质林改造的整地过程中,会对草本层生物多样性造成一定的负面影响,所以在低质林改造的过程中,减少对森林植被的破坏,注意保护稀有植物,进而保护森林生物多样性,有利于森林生态系统最快速恢复稳定,通过改造样地与保留样地的草本层生物多样性指数比值的平均值对比分析,揭草皮子整地方式比暗穴整地方式对草本层植物生物多样性的负面影响小。运用主成分分析方法对试验区内样地的生物多样性进行综合打分的对比评析,揭草皮子整地方式下的改造样地比暗穴整地方式下的改造样地的得分高,说明揭草皮子整地方式优胜于暗穴整地方式,因此在低质林改造整地方式的选择中,揭草皮子整地方式和暗穴整地方式相比,可优先考虑揭草皮子整地方式方法。
为了森林资源可持续的开发利用,在森林采伐利用前,应制定完善的开采与恢复体系,采用合理的采伐方式 避免乱砍滥伐 采伐结束后采用最优更新造林方法,加快森林生态系统的恢复。在森林抚育中,加强对残次树木的清理,有助于改善林内自然环境,有利于幼苗及林下其他植物的生长,加快森林生态系统生物多样性的恢复。
】
[1]邓伦秀,杨荣和.整地方式与直杆桉造林早期效益研究[J].贵州林业科技,1997,25(1):47 -49.
[2]王长庭,龙瑞军,丁路明,等.草地生态系统中物种多样性、群落稳定性和生态系统功能的关系[J].草业科学,2005,22(6):1-7.
[3]赵存玉,王 涛,董治宝,等.科尔沁沙地草场物种多样性特征及其与生境的关系[J].草业科学,2007,25(10):11 -18.
[4]王顺霞,武高林,王占军,等.宁南山区不同造林整地方式对草地植物群落特征的影响[J].草业科学,2009,26(12):87 -91.
[5]李德生,张 萍.论森林生物多样性含义及保护[J].山东林业科技,1996,104(3):28 -30.
[6]刘增文.森林生态系统的物质积累与循环[M].北京:中国林业出版社,2009:118.
[7]杨学春,董希斌,姜 帆,等.黑龙江省伊春林区低质林林分评定[J].东北林业大学学报,2009,37(10):10 -12.
[8]张 泱,姜中珠,董希斌,等.小兴安岭林区低质林类型的界定与评价[J].东北林业大学学报,2009,37(11):99 -102.
[9]刘恩斌.天目山自然保护区生物多样性的熵值测度研究[J].南京林业大学学报(自然科学版),2007,31(5):87-91.
[10]洪 伟,吴承祯.Shannon-Wiener指数的改进[J].福建林学院学报,1999,7(2):120 -124.
[11]何永涛,曹 敏,唐 勇,等.滇中地区常绿阔叶林树种多样性比较研究[J].山地学报,2000,18(4):322 -328.
[12]刘美爽,杨卫东.森林生物多样性可持续发展的探讨[J].林业科技,2006,31(4):66 -68.
[13]贺金生,陈伟烈,李凌浩.中国亚热带东部常绿阔叶林主要类型的群落多样性特征[J].植物生态学报,1988,22(4):303-311.
[14]吴承祯,洪 伟.万木林中亚热带群落物种多样性研究[J].福建林学院学报,1996,16(1):33 -37.
[15]吴晓春,金宏伟,高云波,等.试论森林树种生物多样性保的遗传学理论基础[J].林业科技,2000,25(3):11 -13.
[16]樊后保.格氏栲群落的结构特征[J].林业科学,2000,36(2):6-12.
[17]马建忠,杨桂华,韩明跃,等.梅里雪山国家公园生物多样性保护规划方法研究[J].林业调查规划,2008,33(3):119 -123.
[18]蒋春颖,史明昌.红松混交林重要值、生物多样性与更新的关系研究[J].林业调查规划,2010,35(6):44 -48.
猜你喜欢
中华建设(2024年3期)2024-03-16 08:04:52
中华建设(2023年10期)2023-10-07 05:23:04
建筑与预算(2022年6期)2022-07-08 13:59:18
少儿美术(2020年8期)2020-11-06 05:21:38
海峡姐妹(2019年8期)2019-09-03 01:00:56
海峡姐妹(2018年11期)2018-12-19 05:18:28
东北林业大学学报(2017年10期)2017-10-24 11:50:18
林业科技(2017年4期)2017-08-10 06:02:43
小说月刊(2014年1期)2014-04-23 08:59:59
建筑与预算(2014年5期)2014-04-11 07:29:28
