
食料对大猿叶虫不同地理种群滞育诱导的影响
2011-08-27 00:40:26于洪春邓佳佳薛芳森
植物保护 2011年4期
于洪春, 王 蕾, 邓佳佳, 薛芳森, 张 原
(1.东北农业大学农学院,哈尔滨 150030; 2.江西农业大学昆虫研究所,南昌 330045;3.黑龙江省植检植保站,哈尔滨 150090)
食料对大猿叶虫不同地理种群滞育诱导的影响
于洪春1, 王 蕾1, 邓佳佳1, 薛芳森2, 张 原3
(1.东北农业大学农学院,哈尔滨 150030; 2.江西农业大学昆虫研究所,南昌 330045;3.黑龙江省植检植保站,哈尔滨 150090)
[目的]探求食料对大猿叶虫不同地理种群滞育的影响。[方法]在室内一定的温湿度条件下,观察比较大猿叶虫哈尔滨种群、泰安种群和龙南种群取食白菜、油菜、雪里蕻、萝卜和独行菜5种不同食料的滞育反应。[结果]食料对哈尔滨种群的滞育诱导作用不依赖于光周期,而对泰安种群和龙南种群的滞育诱导作用依赖于光周期。取食白菜、油菜、雪里蕻、萝卜和独行菜5种食料,大猿叶虫哈尔滨种群在25℃的滞育率为89.5%~100%,在28℃的滞育率为76.9%~100%;泰安种群在 25℃的滞育率为21.7%~100%,在28℃的滞育率为4.5%~90.1%;龙南种群在25℃的滞育率为2.5%~100%,在28℃的滞育率为1.1%~90.9%。哈尔滨种群和泰安种群在25℃和28℃取食独行菜滞育率较低;龙南种群在25℃取食萝卜滞育率较低;在28℃取食白菜滞育率较低。[结论]食料对不同地理种群的大猿叶虫滞育诱导的影响显著。
大猿叶虫; 滞育诱导; 地理种群; 食料; 光周期
大猿叶虫(ColaphellusbowringiiBaly)是一种重要的十字花科蔬菜食叶害虫,在国内分布广泛。大猿叶虫具有明显的滞育习性,在我国各地均以成虫在土壤中滞育越冬[1-3]。温度和光周期对大猿叶虫滞育的诱导作用已有研究报道,认为光周期和温度是大猿叶虫滞育诱导的主要因子[4]。食料是影响昆虫滞育诱导的另一个重要因子,为探求食料对大猿叶虫不同地理种群滞育的影响,本文研究了5种不同食料对大猿叶虫不同地理种群的滞育诱导反应。
1 材料与方法
1.1 供试昆虫
大猿叶虫黑龙江哈尔滨种群(45°45′N,126°41′E),采集于哈尔滨市东北农业大学园艺试验站,经成虫繁殖而得;山东泰安种群(36°2′N,117°1′E)和江西龙南种群(24°9′N,114°8′E),由江西农业大学薛芳森教授惠赠的大猿叶虫卵和成虫繁殖而得。
1.2 食料
供试食料均为十字花科植物,白菜(Brassica pekinensis),品种为‘牡丹江二号’,山东省昌邑市鑫昌种子有限公司生产;雪里蕻(Brassica jinceavar.multiceps),国有哈尔滨市种子公司生产;油菜(Brassica napus),品种为‘苏州青油菜’,国有哈尔滨市种子公司生产;萝卜(Raphanus sativus),品种为‘特大红萝卜’,山东省昌邑市鑫昌种子有限公司生产;独行菜(Lepidum sativum),采自东北农业大学园艺试验站野生独行菜。
1.3 试验方法
本试验在哈尔滨东北农业大学昆虫学实验室内进行,设置温度为25℃和28℃,光周期为L∥D=18 h∥6 h、L∥D=16 h∥8 h、L∥D=14 h∥10 h、L∥D=12 h∥12 h、L∥D=10 h∥14 h、L∥D=8 h∥16 h、L∥D=6 h∥18 h,观察和比较大猿叶虫不同地理种群取食白菜、油菜、雪里蕻、萝卜和独行菜5种不同食料的滞育反应。
将不同地理种群的大猿叶虫分别在25℃L∥D=12 h∥12 h的光照培养箱内饲养,待成虫交尾产卵后,将新产的卵分别收集起来进行孵化。收集同批次刚孵化出来的初孵幼虫,用软毛刷转移到放有新鲜食料的养虫盒内,每盒60~70头幼虫,置于所设温度和光周期的光照培养箱中,每处理重复3次。在幼虫接近老熟时,将幼虫分别转移到放入灭菌潮湿细土的新盒中,供老熟幼虫入土化蛹。待成虫羽化出土后,及时将新羽化出土的成虫分别转移到另外的装有灭菌潮湿细土的新盒中,按羽化出土的数量每盒放成虫20~60头。每天更换1次新鲜食料。待成虫进入滞育后,检查并统计入土滞育成虫数,计算滞育率。
滞育判断:参考Xue的判断方法[4]并做适当修改,在羽化出土后16 d检查和统计入土滞育成虫数量。
数据处理与分析:试验数据计算后获得的滞育率,用DPS V3.01专业版软件,经反正弦平方根处理后采用DPS程序包进行方差分析,选用Duncan’s法进行差异显著性比较。
2 结果与分析
大猿叶虫哈尔滨种群、泰安种群和龙南种群取食白菜、油菜、雪里蕻、萝卜和独行菜5种不同食料的滞育反应见图1~3。
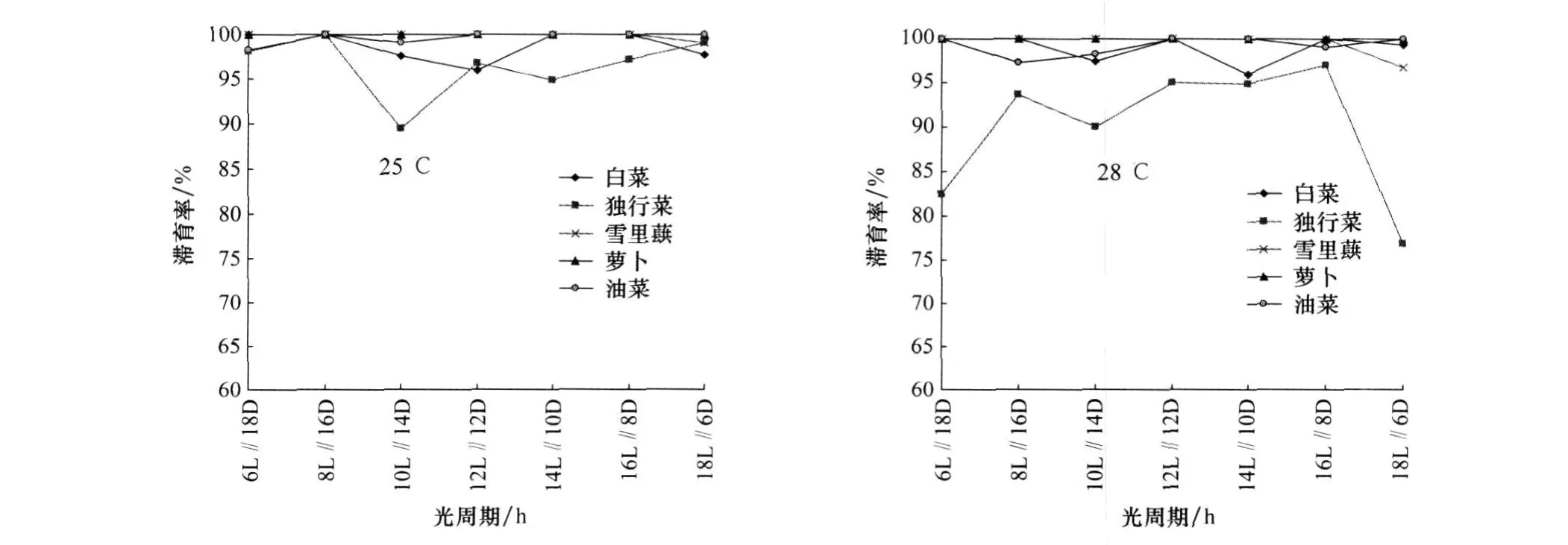
图1 不同食料对大猿叶虫哈尔滨种群在25℃和28℃不同光周期下的滞育诱导
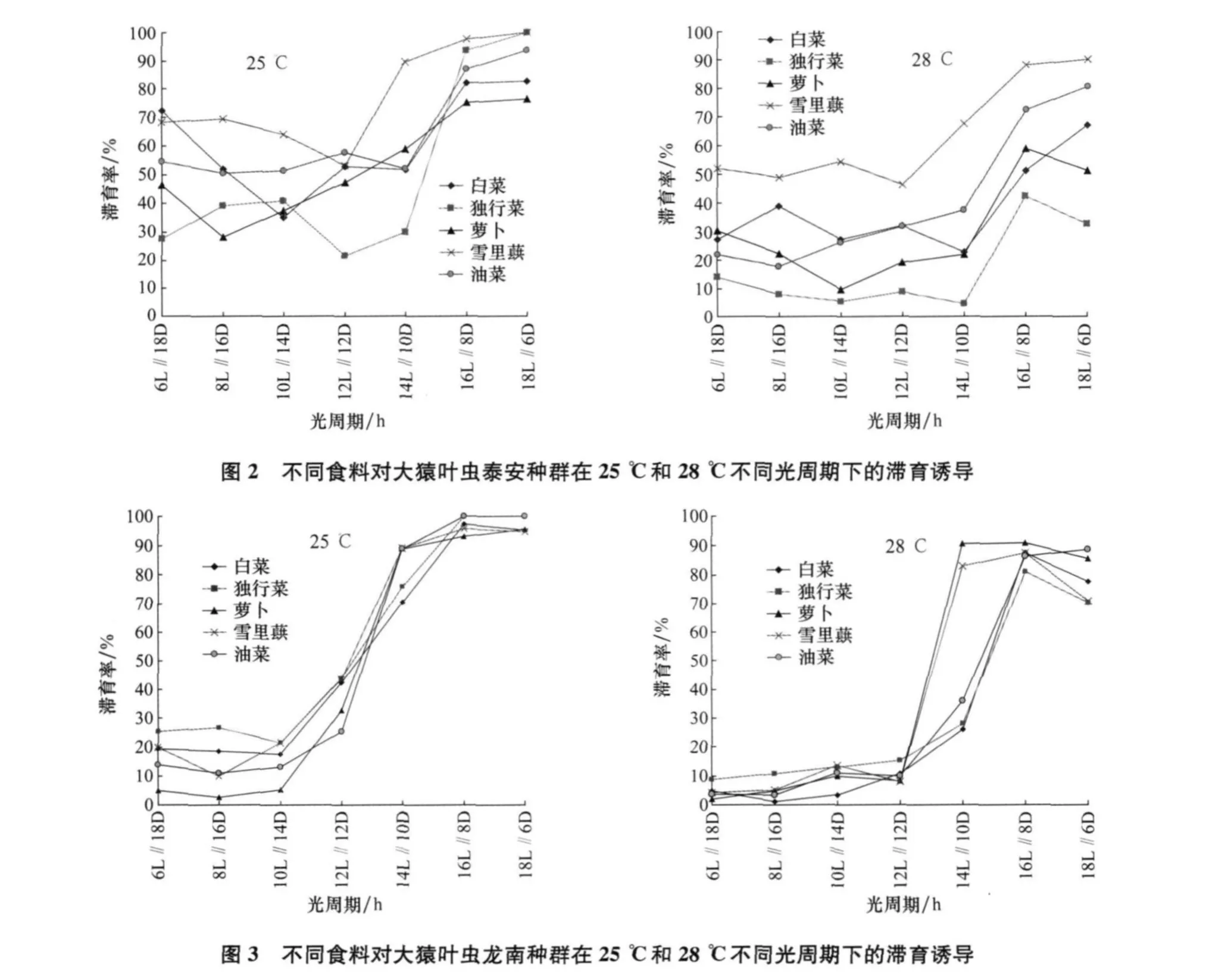
从图1~3可以看出,大猿叶虫哈尔滨种群取食5种食料滞育率均较高,在25℃取食5种食料的滞育率为89.5%~100%;在28℃为76.9%~100%。大猿叶虫泰安种群在25℃取食5种食料的滞育率为21.7%~100%;在28℃为4.5%~90.1%。大猿叶虫龙南种群在25℃取食5种食料的滞育率为2.5%~100%;在28℃为1.1%~90.9%。各地理种群的大猿叶虫食料间Duncan多重比较结果见表1,表明大猿叶虫3个地理种群取食不同寄主植物滞育率存在差异。在25℃和28℃,独行菜对大猿叶虫哈尔滨种群滞育诱导率最低,与其他食料相比差异极显著。泰安种群在25℃取食独行菜和萝卜滞育率较低,在28℃取食独行菜滞育率最低,与其他食料相比差异极显著。龙南种群在25℃取食萝卜滞育率最低,在28℃取食白菜滞育率最低,与其他食料相比差异显著。
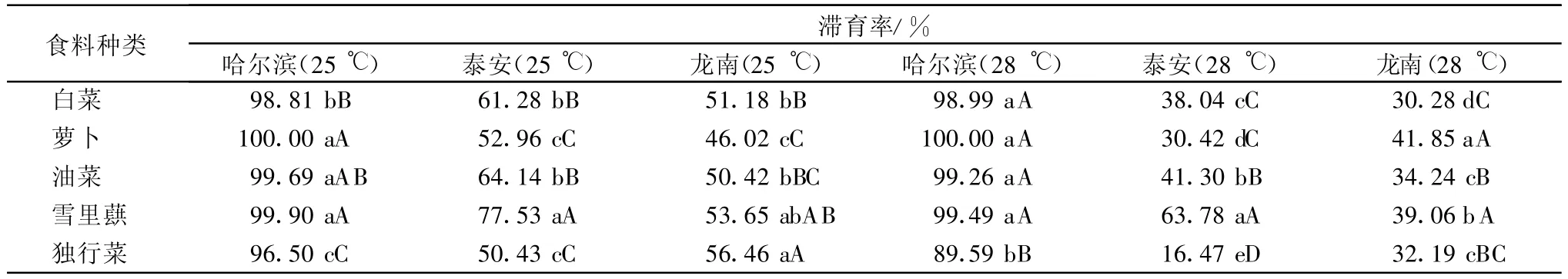
表1 食料种类对大猿叶虫不同地理种群滞育诱导的差异显著性分析1)
经二因子随机设计方差分析和Duncan新复极差法多重比较检验,在25℃,食料对大猿叶虫哈尔滨种群滞育诱导差异极显著(d f=4,F=7.178 82,p=0.000 6<0.01),光周期对其滞育诱导差异不显著(d f=6,F=1.302 22,p=0.294>0.05),食料与光周期组合对其滞育诱导差异极显著(d f=24,F=5.487 77,p<10-5)。在28℃,食料对大猿叶虫哈尔滨种群滞育诱导差异也极显著(d f=4,F=18.801 81,p<10-5),光周期对其滞育诱导差异不显著(df=6,F=1.069 78,p=0.407 63>0.05),食料与光周期组合对其滞育诱导差异极显著(df=24,F=7.090 53,p<10-5)。说明食料和光周期对大猿叶虫哈尔滨种群滞育诱导存在显著的交互作用;但食料的滞育诱导作用不依赖于光周期。大猿叶虫哈尔滨种群在25℃、L∥D=10 h∥14 h光周期下和28℃、L∥D=18 h∥6 h光周期下取食独行菜诱导的滞育发生率最低。
大猿叶虫泰安种群在25℃食料对其滞育诱导差异极显著(df=4,F=6.075 63,p=0.001 59<0.01),光周期对其滞育诱导差异极显著(df=6,F=11.393 37,p<10-5),食料与光周期组合对其滞育诱导差异极显著(df=24,F=14.548 76,p<10-5)。在28℃,食料对其滞育诱导差异也极显著(df=4,F=36.418 59,p<10-5),光周期对其滞育诱导差异极显著(d f=6,F=24.602 27,p<10-5),食料与光周期组合对其滞育诱导差异极显著(df=24,F=8.918 47,p<10-5)。滞育诱导中食料与光周期间存在明显的交互作用,食料的滞育诱导作用依赖于光周期,取食不同食料的大猿叶虫的滞育率随光周期日长的延长而升高。大猿叶虫泰安种群在25℃、L∥D=12 h∥12 h光周期下取食独行菜,以及在L∥D=8 h∥16 h的光周期下取食萝卜,可以获得较低的滞育率;在28℃小于14 h日长的光周期下取食独行菜可以诱导较低的滞育率。
大猿叶虫龙南种群在25℃食料对其滞育诱导差异显著(df=4,F=3.859 25,p=0.014 7<0.05),光周期对其滞育诱导差异极显著(d f=6,F=117.927 86,p<10-5),食料与光周期组合对其滞育诱导差异极显著(d f=24,F=3.856 17,p=0.000 01<0.01)。在28℃,食料对其滞育诱导差异不显著(df=4,F=1.180 2,p=0.344 58>0.05),光周期对其滞育诱导差异极显著(d f=6,F=40.858 26,p<10-5),食料与光周期组合对其滞育诱导差异极显著(df=24,F=12.992 25,p<10-5)。这一结果说明,在高温下,光周期对大猿叶虫龙南种群滞育诱导起了关键作用。滞育诱导中食料与光周期间存在着非常明显的交互作用,食料的滞育诱导作用强烈依赖于光周期,取食不同食料的大猿叶虫的滞育率均随光周期日长的延长而升高,随着光周期中的日长缩短而显著降低。大猿叶虫龙南种群在25℃小于10 h日长的光周期下取食萝卜可以诱导较低的滞育率;在28℃小于12 h日长的光周期下取食白菜滞育率较低。
在25℃和28℃,比较大猿叶虫3个地理种群取食5种食料在L∥D=12 h∥12 h光周期下的滞育诱导效应,其结果见图4。
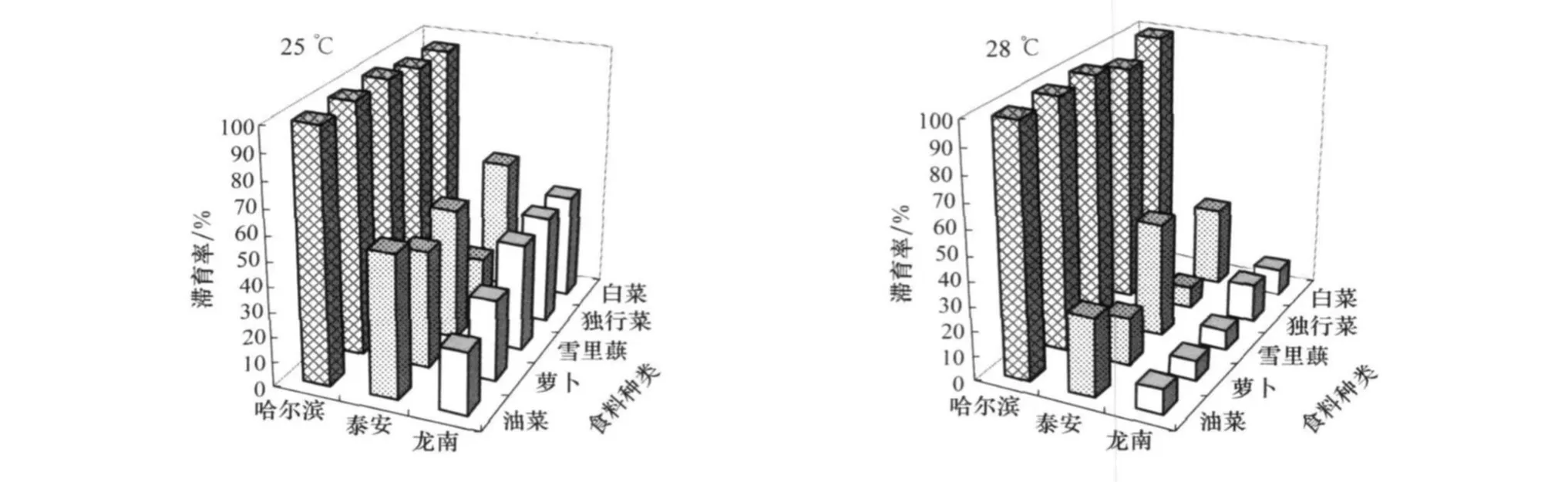
图4 在25℃和28℃,L∥D=12 h∥12 h光周期下食料对不同地理种群大猿叶虫的滞育诱导
经二因子随机设计方差分析和Duncan新复极差法多重比较检验,在25℃和28℃及L∥D=12 h∥12 h光周期下,食料对3个地理种群滞育诱导差异极显著,不同地理种群取食同一寄主食料滞育率差异极显著(25℃,df=2,F=68.214 99,p=0.000 01;28℃,df=2,F=149.490 33,p<10-5)。取食油菜、萝卜、白菜和雪里蕻食料,江西龙南种群滞育率最低,山东泰安地理种群次之,哈尔滨地理种群最高;取食独行菜,山东泰安种群滞育率最低,江西龙南种群次之,哈尔滨种群最高。
3 讨论
食物(寄主)种类影响昆虫的滞育诱导已经有较多报道。在林间,玫瑰色卷蛾(Choristoneura rosaceana)幼虫取食红花槭(Acer rubrum)、黑白蜡树(Fraxinus nigra)、北美白桦(Betula papyri f era)和弗吉尼亚李(Prunusvirginiana)4种不同的食料植物,进入滞育的比例不同;幼虫取食弗吉尼亚李继续发育产生第2代,而取食北美白桦会诱导中等量的滞育[5]。作者研究表明,食料种类不仅对同一地理种群的大猿叶虫滞育诱导差异显著,对不同地理种群的大猿叶虫滞育诱导差异也显著,不同地理种群的大猿叶虫取食同一寄主食料,其滞育率有显著差异,表明大猿叶虫的滞育存在着地理变异。总体上看,在短日照光周期下,取食同一寄主食料,大猿叶虫黑龙江哈尔滨种群的滞育率大于山东泰安种群的滞育率,山东泰安种群的滞育率大于江西龙南种群的滞育率,表明昆虫的滞育随纬度而变化,高纬度地理种群比低纬度地理种群有更高的滞育率。昆虫的滞育随纬度而变化已有报道,斑足大眼长蝽(Geocoris punctipes)的滞育率在短于L14:D10 h的光周期下,Kentucky种群(38°04′N)的滞育率均比Georgia种群(31°28′N)的高,Georgia种群在任一短光周期下进入滞育不超过50%,而Kentucky种群在短光照条件下可有80%以上进入滞育[6]。大猿叶虫滞育诱导中食料与光周期间存在着非常明显的交互作用,但食料对大猿叶虫不同地理种群的滞育诱导作用不同,哈尔滨种群的滞育诱导作用不依赖于光周期,而泰安种群和龙南种群的滞育诱导作用依赖于光周期,具有明显的光周期反应,尤以龙南种群更为显著,表明食料虽是大猿叶虫滞育诱导的一个因子,但其诱导作用与光周期关系密切,光周期仍是滞育诱导的更重要因子。这些研究结果有助于我们探明大猿叶虫生活史和滞育多型现象形成的原因。
[1]薛芳森,李爱青,朱杏芬,等.大猿叶虫生活史的研究[J].昆虫学报,2002,45(4):494-498.
[2]董宇奎,周昱晨,李照会,等.大猿叶虫山东种群生物学特性研究[J].植物保护,2007,33(2):70-73.
[3]胡志凤,于洪春,孙文鹏,等.哈尔滨地区大猿叶虫发育历期与生物学特性[J].昆虫知识,2008,45(6):909-912.
[4]Xue F S,Spieth H R,Li A Q,et al.The role of photoperiod and temperature in determination of summer and winter diapause in the cabbage beetle,Colaphellusbowringi(Coleoptera:Chrysomelidae)[J].J Insect Physiol,2002,48:279-286.
[5]Hunter M D,McNeil J N.Host-plant quality influences diapause and voltinism in a poly phagous insect herbivore[J].Ecology,1997,78:977-986.
[6]John R R,Kenneth V Y,Blake L N.Variation in diapause response between geographical populations of the predatorGeocoris punctipes(Heteroptera:Geocoridae)[J].Ann Entomol Soc Am,2001,94(1):116-122.
Effects of different foods on diapause induction in three geographic populations of Colaphellus bowringii
Yu Hongchun1, Wang Lei1, Deng Jiajia1, Xue Fangsen2, Zhang Yuan3
(1.College of Agriculture,Northeast Agricultural University,Harbin150030,China;2.Institute of Entomology,Jiangxi Agricultural University,Nanchang330045,China;3.Heilongjiang Province Plant Quarantine and Protection Station,Harbin150090,China)
[Objective]The effects of food on diapause induction in different geographic populations ofColaphellus bowringiiBaly was investigated.[Method]The diapause reaction of Harbin,Tai’an and Longnan populations ofC.bowringii,fed withBrassica pekinensis,B.napus,B.jincea,Raphanus sativusandLepidum sativum,were observed at constant temperature and humidity in the laboratory.[Result]The diapause induction of food in Harbin population was not related to photoperiod,but Tai’an and Longnan populations were significantly related to photoperiod.Fed withB.pekinensis,B.napus,B.jincea,R.sativusandL.sativum,the percentage of diapaused individuals was 89.5%-100%at 25℃and 76.9%-100%at 28℃in Harbin population,but the diapause rate was 21.7%-100%at 25 ℃and 4.5%-90.1%at 28 ℃in Tai’an population,and 2.5%-100%at 25℃and 1.1%-90.9%at 28℃in Longnan population.When fed with the leaves ofL.sativum,the percentage of diapaused individuals was lower in Harbin and Tai’an populations at 25℃ and 28℃.However,the diapause rate was lower at 25℃when fed with the leaves ofR.sativusand was lower at 28℃when fed with the leaves ofB.pekinensisin Longnan population.[Conclusion]Different foods had different effects on diapause induction in the three populations.
Colaphellus bowringii; diapause induction; geographic population; food; photoperiod
S 433.5
A
10.3969/j.issn.0529-1542.2011.04.013
2010-06-22
2010-08-16
国家自然科学基金项目(30460074)
联系方式 Tel:0451-55191053;E-mail:hongcyu@yahoo.com.cn
猜你喜欢
遗传(2023年9期)2023-09-25 09:31:44
农业科技通讯(2023年1期)2023-02-12 07:08:50
广东蚕业(2022年8期)2022-09-30 01:51:00
中国饲料(2019年8期)2019-06-14 06:25:32
疯狂英语·新悦读(2019年5期)2019-05-15 08:53:56
疯狂英语·新悦读(2019年5期)2019-05-15 08:53:50
少儿科学周刊·少年版(2017年7期)2017-09-29 08:02:41
少儿科学周刊·儿童版(2017年7期)2017-09-29 02:22:58
福建农业科技(2016年4期)2016-08-11 01:32:58
作文大王·低年级(2015年11期)2015-05-30 10:48:04
