
呋喃虫酰肼对舞毒蛾解毒酶活力的影响1)
2011-08-09 11:07:58严善春曹传旺
东北林业大学学报 2011年9期
李 慧 严善春 曹传旺
(东北林业大学,哈尔滨,150040)
舞毒蛾(Lymantria dispar)是我国重要的森林害虫,同时又是国际上重要的检疫性有害生物,严重影响木材外贸出口[1-3]。目前,化学药剂防治仍然是国内外对舞毒蛾的主要控制措施。呋喃虫酰肼(JS118)是我国具有知识产权的一种高效促蜕皮仿生杀虫剂[4-5],其主要利用昆虫生长过程中产生的激素物质,干扰昆虫正常的生长发育。研究表明,呋喃虫酰肼对鳞翅目害虫如甜菜夜蛾、斜纹夜蛾、小菜蛾、二化螟等都表现出较高的生物活性,同时对非靶标生物有很高的安全性,具有低药量、持续长、稳定、高效,对人畜安全等特点[6-7]。目前,尚未有将呋喃虫酰肼用于防治林木害虫的相关报道。
昆虫解毒酶主要包括羧酸酯酶(CarE)、谷胱甘肽S-转移酶(GSTs)和多功能氧化酶O-脱甲基(MFOs)[8-9]。磷酸酯酶分为酸性磷酸酯酶(ACP)和碱性磷酸酯酶(ALP),是一种与昆虫抗性有关的重要水解酶[10]。昆虫解毒酶能被各种外源化合物诱导,使昆虫在受到严重的化学环境压力作用时迅速做出反应,从而存活下来[11]。因此昆虫的解毒酶在杀虫剂代谢、昆虫抗药性等方面都具有重要作用。本研究以舞毒蛾幼虫为对象,从生化水平探讨了亚致死浓度呋喃虫酰肼对舞毒蛾的生物活性及对其幼虫体内解毒酶活性的影响,为应用该药剂对舞毒蛾的防治研究提供理论依据。
1 材料与方法
1.1 试虫与饲养
2010年3月在东北林业大学校园及其示范林场内采集舞毒蛾卵块。将采回的虫卵置于恒温培养箱内孵化,幼虫用人工饲料饲养,每天观察并及时更换饲料,定期清理幼虫的排泄物和食物残渣。恒温饲养条件:温度(25±1)℃,相对湿度75%,16 h光照,8 h黑暗。
1.2 供试药剂
10%呋喃虫酰肼悬浮剂(JS118)购自江苏省农药研究所有限公司;α-乙酸萘酯、硝基苯基磷酸二钠、巴比妥钠、考马斯亮蓝G-250、血清白蛋白、乙二胺四乙酸均购自国药集团化学试剂有限公司;还原型谷胱甘肽、1-氯-2,4-二硝基苯、对硝基苯甲醚、毒扁豆碱、固蓝B盐、十二烷基硫酸钠、苯甲基磺酰氟、二硫苏糖醇、硫脲均购自美国Sigma公司;还原型辅酶Ⅱ四钠购自瑞士Rocha公司;磷酸氢二钠和磷酸二氢钠及其他试剂均为分析纯。
1.3 毒力测定
毒力测定采用叶片药膜法[12-13]。用蒸馏水将10%呋喃虫酰肼等比稀释6个梯度,其中2、3龄幼虫选用的质量浓度梯度为 1.00、0.75、0.50、0.25、0.10、0.05、0.01 mg·L-1;4、5、6 龄幼虫选用的质量浓度梯度为 1.50、1.25、1.00、0.75、0.50、0.25、0.10 mg·L-1,以蒸馏水为对照。将未接触过任何药剂的白桦(Betula platyphylla Suk.)叶片浸入稀释好的药液中,10 s后取出,置于阴凉处晾干。用蘸有充足水分的脱脂棉裹住叶柄,放入透气性良好的塑料培养皿(直径d=9 cm)中。将健康、大小一致的2~6龄舞毒蛾幼虫接到白桦叶片上,每龄每皿放入10头,每龄每个质量浓度重复3次。置于恒温培养箱内饲养,24 h后检查死亡数,以毛笔轻触幼虫不能活动视为死亡。
1.4 亚致死浓度处理
根据毒力测定结果,用选定24 h的LC50剂量呋喃虫酰肼处理白桦叶片,饲喂健康、大小一致,且饥饿12 h的3龄和5龄舞毒蛾幼虫,每龄每皿放入10头,处理方法和饲养条件同1.3,以蒸馏水处理作对照。分别于处理 6、12、24、36、48 h 后,挑选活泼的3龄和5龄舞毒蛾幼虫,于冰箱中-80℃储存,用于酶活性测定。
1.5 解毒酶活性测定
羧酸酯酶(CarE)活性:参照 Van Asperen[14]的方法略有改动。在试管中分别加入3×10-4mol·L-1的α-乙酸萘酯底物(含1∶1 毒扁豆碱)5.0 mL,0.04 mol·L-1、pH 值7.0 磷酸缓冲液0.8 mL,酶液0.2 mL,立即混匀,在30℃水浴振荡条件下反应30 min,然后每支试管中加入显色剂1.0 mL,室温下静置30 min,于600 nm处测定OD值。以酶量(mL)为自变量,OD值为因变量,根据羧酸酯酶标准曲线计算出每毫升酶液生成的α-萘酚量,再根据蛋白质质量(mg),计算出羧酸酯酶的比活力。酶活力单位以mol·g-1·min-1表示。
磷酸酯酶(ACP和 ALP)活性:参照Bessey等[15]的方法略作修改。ACP活性测定采用对硝基苯磷酸二钠做底物,反应混合物为 0.2 mol·L-1、pH 值4.6 醋酸缓冲液2.3 mL,7.5 ×10-3mol·L-1对硝基苯磷酸二钠溶液0.5 mL和0.1 mL酶液。混匀30℃水浴反应15 min后加入0.1 mol·L-1的氢氧化钠2.0 mL终止反应,混匀后于400 nm处测定OD值。对照中用缓冲液代替酶液。ALP活性测定,用0.4 mol·L-1,pH值9.6巴比妥钠—盐酸缓冲液代替0.2 mol·L-1、pH值4.6的醋酸缓冲液,其余方法同ACP活性测定。酶活力以mol·g-1·min-1表示。
谷胱甘肽S-转移酶(GSTs)活性:参照Booth[16]的方法。在试管中分别加入0.066 mol·L-1、pH值7.0的磷酸缓冲液2.5 mL,0.5 mol·L-1GSH 0.3 mL,0.03 mol·L-1CDNB 0.1 mL,酶液0.1 mL,立即混匀,在27℃,于340 nm处测定5 min内OD的变化值。酶活力单位以ΔOD·mg-1·min-1表示。
多功能氧化酶(MFOs)活性:参考Hansen等[17]方法略作修改。取 4 ×10-3mol·L-1对硝基苯甲醚 1.0 mL,0.1 mol·L-1、pH 值7.8 磷酸缓冲液0.8 mL,5 ×10-4mol·L-1NADPH-Na40.2 mL 和酶液 1.0 mL,以等体积的 0.1 mol·L-1、pH值7.8磷酸缓冲液作对照,摇匀置于37℃水浴中反应30 min,然后在反应体系中加入1.0 mol·L-1盐酸1.0 mL终止反应,再加入5.0 mL氯仿萃取,静止10 min后,在氯仿层移取3.0 mL 到另一组试管内,再加入 0.5 mol·L-1氢氧化钠 3.0 mL溶液萃取,静止10 min,取水相2.0 mL于比色皿中,于400 nm处测定OD值。酶活力单位以mol·g-1·min-1表示。
1.6 蛋白质质量浓度测定
采用Bradford考马斯亮蓝G-250法[18]。
1.7 低剂量呋喃虫酰肼对舞毒蛾各发育指标的影响测定
采用叶片药膜法,取用LC10和LC30剂量呋喃虫酰肼处理的白桦叶片,饲喂刚蜕皮大小一致的3龄舞毒蛾幼虫,每培养皿(直径9 cm)1头,置于恒温培养箱内饲养,以清水处理为对照。在其整个幼虫发育期,均用LC10和LC30剂量呋喃虫酰肼处理的叶片喂养,待幼虫羽化为成虫后,以5%的新鲜蜜糖水继续喂养。每10头试虫为一组,重复3次。从4龄开始,每隔12 h观察并统计各龄期的成活率、发育历期、化蛹率、羽化率等发育指标,直到成虫死亡。
1.8 数据统计分析
利用POLO软件处理分析毒力测定结果,计算致死中浓度(LC50)和低浓度(LC10和LC30)及其95%置信区间;采用SPSS17.0软件进行方差分析,差异显著性检验采用LSD法,显著水平 α =0.05或 α=0.01。
2 结果与分析
2.1 呋喃虫酰肼对舞毒蛾幼虫的毒力
从表1可以看出,呋喃虫酰肼对不同龄期的舞毒蛾幼虫均表现出较高的活性,虫龄越小,对药剂越敏感,随着幼虫龄期的增长,所需药量逐渐增加。选择处理24 h的LC50作为试验剂量处理3龄和5龄舞毒蛾幼虫(LC50分别为0.369、0.842 mg·L-1),测定呋喃虫酰肼对其体内解毒酶活性的影响。并选择舞毒蛾幼虫24 h的低浓度LC10和LC30饲喂大龄幼虫研究呋喃虫酰肼对其生长发育的影响。

表1 呋喃虫酰肼对舞毒蛾各龄幼虫的毒性
2.2 呋喃虫酰肼对3龄舞毒蛾幼虫解毒酶活性的影响
呋喃虫酰肼对舞毒蛾3龄幼虫体内解毒酶活性影响明显。从表2可以看出,呋喃虫酰肼对3龄幼虫CarE活性具有先激活后抑制作用,处理6~12 h,CarE活性明显被激活,其中处理6 h时,对CarE的激活作用最强,为同期对照的1.243倍,差异极显著(p<0.01);24 h后CarE活性逐渐被抑制,并随处理时间的延长,对该酶活抑制作用逐渐增强。处理过程中ACP活性明显被抑制,并随处理时间的延长,酶活抑制作用增强;ALP活性则表现为先抑制后激活再抑制作用,处理12 h,ALP活性明显被激活,为同期对照组的1.286倍,差异极显著(p<0.01);24 h后,ALP活性又逐渐被抑制。

表2 呋喃虫酰肼对3龄舞毒蛾幼虫CarE、ACP和ALP活性的影响
从表3中可以看出,呋喃虫酰肼对3龄舞毒蛾幼虫体内GSTs活性具有先激活后抑制作用,处理6 h后,处理组GSTs活性明显高于对照组,为对照组的1.16倍,差异显著(p<0.05);12 h后,GSTs活性逐渐被抑制,且处理时间越长,对该酶活的抑制作用越强,与对照组相比差异均极显著(p<0.01)。对MFOs活性表现为先抑制后激活,处理6~12 h,MFOs活性被抑制,其中处理6 h时,对MFOs的活性抑制作用最强,为同期对照的0.843倍,差异极显著(p<0.01);从24 h起,呋喃虫酰肼对MFOs活性具有明显的激活作用,其中处理36 h时,MFOs激活作用最强,为同期对照的1.393倍,差异极显著(p<0.01),随处理时间的延长,药剂对该酶活激活作用呈下降趋势。说明呋喃虫酰肼对3龄舞毒蛾幼虫体内解毒酶活性的影响比较复杂,对酶活性表现为诱导或抑制作用。
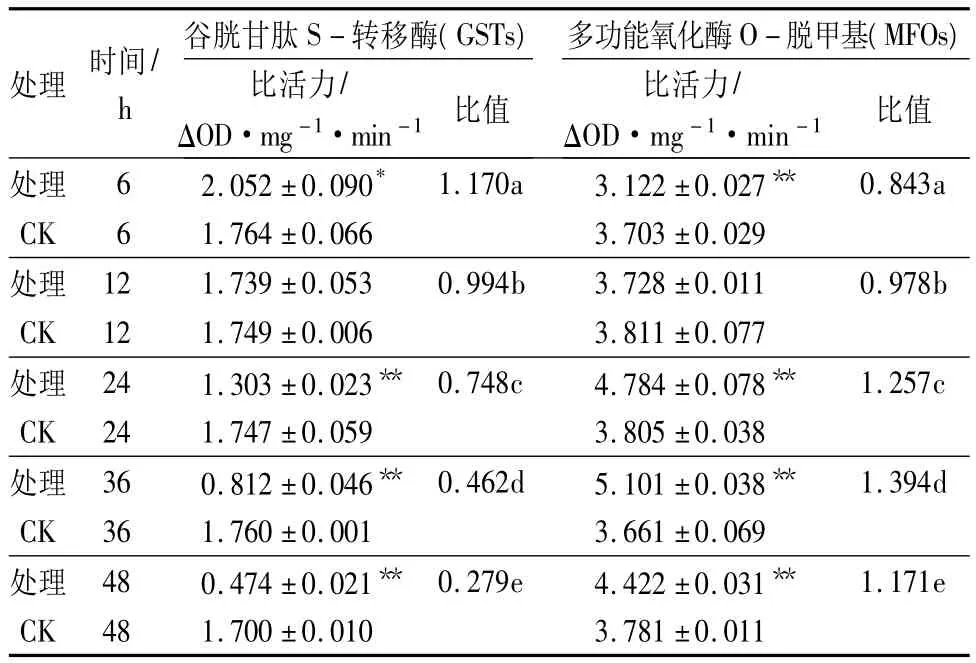
表3 呋喃虫酰肼对3龄舞毒蛾幼虫GSTs和MFOs活性的影响
2.3 呋喃虫酰肼对5龄舞毒蛾幼虫解毒酶活性的影响
表4和表5为呋喃虫酰肼对舞毒蛾5龄幼虫体内解毒酶活性的影响。从表4中可以看出,5龄舞毒蛾幼虫经呋喃虫酰肼处理后,6~12 h,CarE活性明显被抑制,12 h后酶活性逐渐被激活,其中处理24 h时,处理组CarE活性明显高于对照组,为同期对照组的 1.349 倍,差异极显著(p<0.01),36 h 后CarE活性又逐渐被抑制。对ACP活性表现为明显的抑制作用,且随处理时间的延长,对ACP活性的抑制作用增强。对ALP活性表现为先激活后抑制,6 h时ALP活性被激活,为同期对照组的1.106 倍,差异显著(p<0.05),24~48 h,ALP 活性随着处理时间的延长而逐渐被抑制,且差异极显著(p<0.01)。
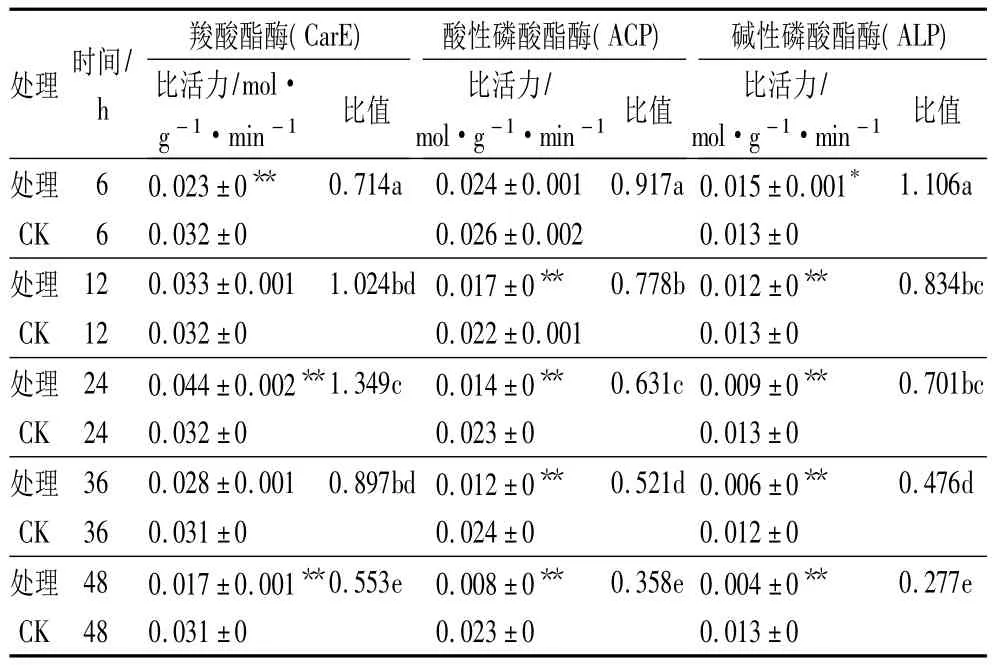
表4 呋喃虫酰肼对5龄舞毒蛾幼虫CarE、ACP和ALP活性的影响
从表5可以看出,呋喃虫酰肼对5龄舞毒蛾幼虫体内GSTs和MFOs存在明显影响。呋喃虫酰肼对5龄舞毒蛾幼虫体内GSTs活性具有明显的抑制作用,且药剂处理时间越长,对GSTs活性抑制作用越强,均表现出差异显著或极显著(p<0.05或p<0.01)。对MFOs活性表现出先激活后抑制,24~36 h,处理组MFOs活性被激活,其中24 h时MFOs的激活作用最强,为同期对照组的1.387倍,差异极显著(p<0.01),48 h时,MFOs活性开始被抑制。
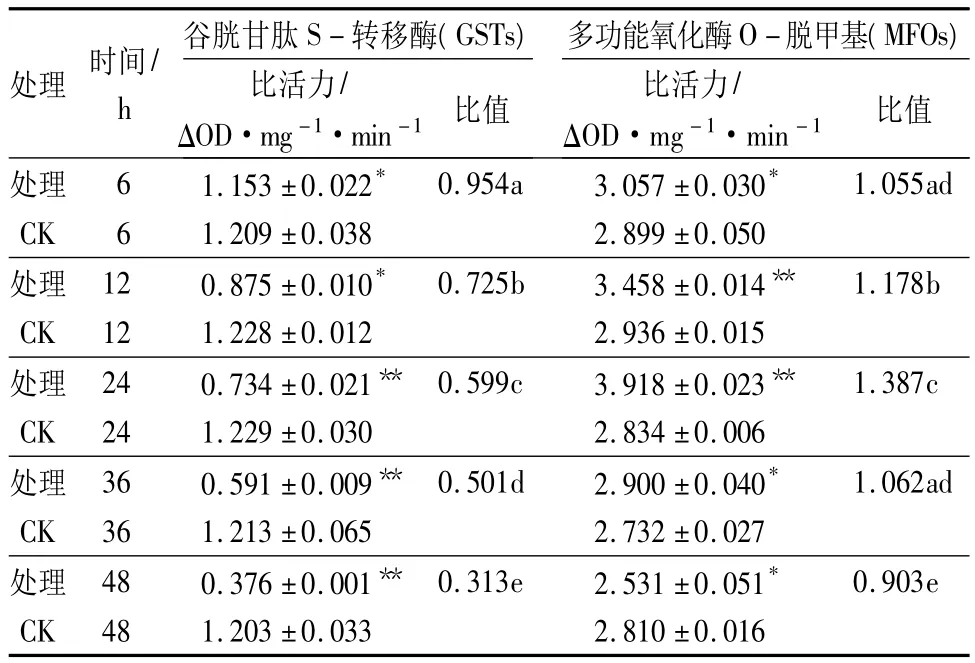
表5 呋喃虫酰肼对5龄舞毒蛾幼虫GSTs和MFOs活性的影响
2.4 低剂量呋喃虫酰肼对舞毒蛾生长发育的影响
由表6可以看出,呋喃虫酰肼对舞毒蛾的生长发育具有明显的影响。与对照相比,LC10和LC30剂量的呋喃虫酰肼均明显的延长了舞毒蛾幼虫的发育时间,随着呋喃虫酰肼质量浓度的增大,幼虫的成活率明显降低,且药剂处理时间越长,幼虫的成活率越低(p<0.05)。
3 结论与讨论
有研究表明,呋喃虫酰肼对甜菜夜蛾的防效明显优于高效氯氰菊酯乳油[19]。李慧等[20]经毒力测定发现,有机磷杀虫剂毒死蜱对舞毒蛾3龄幼虫在24 h的致死中浓度为5.86 mg·L-1,远高于昆虫生长调节剂类杀虫剂呋喃虫酰肼对舞毒蛾3龄幼虫的致死中浓度0.369 mg·L-1,进一步证明呋喃虫酰肼的低毒作用。
非甾醇蜕皮激素类杀虫剂在昆虫体内的解毒代谢与GSTs和 MFOs有关[21];周利琳等[22]研究表明虫酰肼能够诱导CarE和GSTs的活性增强。马志卿等[23]研究表明,用沾有脱氧鬼臼毒素的小麦叶碟饲喂粘虫后,明显抑制其ACP活性,而ALP活性则表现为先诱导后抑制的变化过程。李晓涛等[24]研究发现亚致死剂量氟虫腈对二化螟4龄幼虫体内MFOs活性表现出显著的诱导效应。本研究表明呋喃虫酰肼能有效影响舞毒蛾3龄和5龄幼虫体内解毒酶活性,处理初期,呋喃虫酰肼刺激幼虫使其几种解毒酶活性被逐渐诱导激活,增强水解代谢以达到解毒的目的。随着药剂处理时间的延长,幼虫体内的呋喃虫酰肼含量逐渐增加,解毒酶活性抑制趋势逐渐增强,最终解毒酶活性逐渐被抑制,从而降低了幼虫对该药剂的解毒作用,导致中毒后幼虫本身的生理机能下降、死亡。

表6 低剂量呋喃虫酰肼对舞毒蛾发育指标的影响
经呋喃虫酰肼处理后,舞毒蛾幼虫出现虫体发黑、头壳凸起、无法翻身、身体缩小等中毒症状,且在48 h时几种解毒酶的活性均被抑制,这表明舞毒蛾幼虫体内的解毒作用被中断。低剂量呋喃虫酰肼能够显著影响舞毒蛾幼虫的发育速率和成活率,药剂连续处理15~20 d,幼虫全部死亡,化蛹前幼虫死亡率已达100%。这说明呋喃虫酰肼能明显干扰舞毒蛾的生长发育,对低龄和老龄的舞毒蛾幼虫均具有较高的毒杀作用。因此,呋喃虫酰肼可用作高毒杀虫剂的替代品[6],可作为防治舞毒蛾的安全、有效药剂。
[1]王清树,慈维华.舞毒蛾的发生与防治技术[J].现代化农业,2007(7):44-45.
[2]Kostic'M,Popovic'Z,Brkic'D,et al.Larvicidal and antifeedant activity of some plant-derived compounds to Lymantria dispar L.(Lepidoptera:Limantriidae)[J].Bioresource Technology,2008,99(16):7897-7901.
[3]李景奎,戚大伟.电磁辐射对舞毒蛾卵孵化率的影响[J].中国森林病虫,2007,26(2):9-11.
[4]Hu J Y,Deng Z B,Qin D M.Determination of diacylhydrazinestype insect growth regulator JS-118 residues in cabbage and soil by high performance liquid chromatography with DAD detection[J].Bulletin of Environmental Contamination and Toxicology,2009,83(6):803-807.
[5]Hu J Y,Liu C,Zhang Y C,et al.Hydrolysis and photolysis of diacylhydrazines-type insect growth regulator js-118 in aqueous solutions under abiotic conditions[J].Bulletin of Environmental Contamination and Toxicology,2009,82(5):610-615.
[6]张湘宁.新型昆虫生长调节剂:呋喃虫酰肼[J].世界农药,2005,27(4):48-49.
[7]Guo Cong,Li Dahui,Chen Jinhui,et al.Degradation of furan tebufenozide in laboratory and field trials[J].Science China Chemistry,2010,53(18):1818-1824.
[8]丁双阳,李怀业,李学锋,等.转Bt基因杨树对美国白蛾幼虫中肠解毒酶及乙酰胆碱酯酶的影响[J].东北林业大学学报,2001,29(3):28-30.
[9]Yang Xuemei,Maegolies D C,Zhu Kunyan et al.Host plant-induced changes in detoxification enzymes and susceptibility to pesticides in the twospotted spider mite(Acari:Tetranychidae) [J].Journal of Economic Entomology,2001,94(2):381-387.
[10]马红梅,陈海婴,柳小青,等.德国小蠊磷酸酯酶及谷胱甘肽S-转移酶生化特性的变化与抗药性的关系研究[J].中国媒介生物学及控制杂志,2008,19(5):422-425.
[11]Mohan M,Gujar G T.Local variation in susceptibility of the diamondback moth,Plutella xylostella(Linnaeus)to insecticides and role of detoxification enzymes[J].Crop Protection,2003,22(3):495-504.
[12]马惠,王开运,夏晓明,等.5种新型杀虫剂对甜菜夜蛾幼虫的毒力测定[J].现代农药,2006,5(3):44-46.
[13]Zhao JZ,Li Y X,Collins H L,et al.Monitoring and characterization of diamondback moth(Lepidoptera:Plutellidae)resistance to spinosad[J].Journal of Economic Entomology,2002,95(2):430-436.
[14]Van Asperen K.A study of housefly esterases by means of a sensitive colorimetric method[J].Journal of Insect Physiology,1962,8(4):415-416.
[15]Bessey O A,Lowry O H,Brock M J.A method for the rapid determination of alkaline phosphatase with five cubic millimeters of serum[J].The Journal of Biological Chemistry,1946,164:321-329.
[16]Booth J,Boyland E,Sims P.An enzyme from rat liver catalyzing conjugations with gluthatioze[J].Biochemistry Journal,1961,79(3):516-524.
[17]Hansen L G,Hodgson E.Biochemical characteristics of insect microsomes:N-and O-demethylation[J].Biochemical Pharmacology,1971,20(7):1569-1578.
[18]Bradford M M.A rapid and sensitive method for the quantitation of microgram quantities of protein unitizing the principle of proteindye binding[J].Analytical Biochemistry,1976,72:248-254.
[19]倪珏萍,朱丽梅,侯华民,等.新杀虫剂JS118生物活性研究[J].现代农药,2002,1(1):16-20.
[20]李慧,严善春,王志英,等.毒死蜱对舞毒蛾食物利用和解毒酶及 AChE 活性的影响[J].林业科学研究,2011,24(3):345-349.
[21]黄琳瑞.甜菜夜蛾对虫酰肼抗药性选育和抗药性机理初探[D].北京:中国农业大学,2005.
[22]周利琳,司升云,汪钟信,等.甜菜夜蛾对虫酰肼抗性的生化机制[J].昆虫学报,2009,52(4):386-394.
[23]马志卿,李广泽,冯俊涛,等.脱氧鬼臼毒素对粘虫几种代谢酶系的影响[J].昆虫学报,2007,50(2):186-190.
[24]李晓涛,黄青春,唐振华.氟虫腈对二化螟生长发育的影响及对解毒酶的诱导效应[J].农药学学报,2006,8(3):250-254.
猜你喜欢
四川蚕业(2021年4期)2021-03-08 02:59:58
中国科技纵横(2021年24期)2021-03-02 06:42:52
四川蚕业(2021年3期)2021-02-12 02:38:40
中成药(2018年6期)2018-07-11 03:01:12
林业与生态(2016年8期)2016-12-14 10:55:02
现代农业(2016年6期)2016-02-28 18:42:52
烟草科技(2015年8期)2015-12-20 08:27:14
无机化学学报(2014年6期)2014-02-28 17:32:06
山西大同大学学报(自然科学版)(2013年5期)2013-09-13 10:44:14
食品科学(2013年24期)2013-03-11 18:30:31
