
农杆菌介导P2300-pa-8e抗虫基因转化马铃薯的研究
2011-08-07 09:42杨志超张丽莉卢翠华姜丽丽张正国林忠平胡鸢雷
中国蔬菜 2011年18期
杨志超 张丽莉 卢翠华* 邸 宏 姜丽丽 张正国 林忠平胡鸢雷
(1东北农业大学农学院,黑龙江哈尔滨 150030;2北京大学生命科学学院,北京 100871)
马铃薯(Solanum tuberosumL.)是茄科茄属双子叶植物,是世界四大粮食作物之一,由于它具有适应性强、产量高、营养成分全和加工产业链长等特点,受到了全世界的广泛重视。我国是世界马铃薯生产第一大国,面积和总产量占全世界的1/4(卢翠华 等,2009)。
虫害是造成马铃薯减产的主要原因,其中金龟子科害虫金龟子(幼虫俗称蛴螬)是世界范围内广泛分布的地下害虫之一。金龟子成虫、幼虫均可危害马铃薯,但幼虫危害较大。传统化学药剂防治不仅危害人类健康,而且造成环境污染(姚庆学 等,2003)。自1996年转基因作物商业化以来,全球种植的抗虫转基因作物面积已经达到2 625万hm2(James,2008)。其中对金龟子科害虫具有杀虫作用的苏云金芽胞杆菌cry类基因的相继发现和克隆,为害虫的防治提供了新的途径。Yu等(2006)从Bt185中克隆了P2300-pa-8ea1基因。
本试验将来源于Bt菌株Bt185的P2300-pa-8e基因导入马铃薯中,期望获得抗虫的马铃薯材料,为马铃薯抗性育种提供新资源。
1 材料与方法
1.1 试验材料和菌株
试验于2009年7月~2010年5月在东北农业大学马铃薯实验室进行。
供试马铃薯品种为早大白和克新18号(东北农业大学提供)。
供试农杆菌菌种为LBA4404(北京大学生命科学学院提供),植物表达载体质粒P2300-Pa-8e(图1),利用Patatin启动子驱动P2300-pa-8e(500 bp)基因的表达,质粒由北京大学生命科学学院林中平教授提供。
1.2 试验方法
1.2.1 工程菌液的制备 挑取含有质粒P2300-Pa-8e的农杆菌LBA4404单菌落接种于含有卡那霉素(Kan)、链霉素(Str)和利福平(Rif)的YEB培养基中,避光震荡培养。从中取500 μL菌液接种于50 mL相同的YEB培养基中,以相同条件震荡培养。到对数生长期,在4 000 r·min-1条件下离心5 min,弃上清液,重悬于MS液体培养基中,为工程菌液。
1.2.2 薯块再生培养基的筛选 将马铃薯早大白和克新18号脱毒微型薯灭菌、去皮,切成薄片,放入工程菌液中进行定时侵染,转入共培养培养基,28 ℃黑暗条件下培养 2~3 d。分别接种于9种再生培养基上(表1),诱导薯块出芽。待芽长到2~3 cm时剪下,转入生根培养基生根(丛培琳 等,2008;姜丽丽 等,2009)。
1.2.3 共培养时间对遗传转化的影响 在菌液OD600值为0.5、浸染时间为5 min的条件下,用农杆菌感染薯片,置于 28 ℃黑暗条件下分别培养1、2、3、4、5 d,在诱导培养基上培养20 d,统计抗性愈伤的诱导率。
1.2.4 PCR检测 使用索来宝公司的植物总DNA提取试剂盒提取马铃薯植株的总DNA,以转化植株的总DNA为模板,未转化植株的总DNA作阴性对照,以P2300-pa-8e质粒为阳性对照,根据P2300-pa-8e基因序列,设计特异性引物(PrimerⅠ:5′-GGCGGCAGCATTCAAACTCAA-3′,PrimerⅡ:5′-ATCTCCACCAAGATAGTGTCC-3′),对目标基因进行扩增。PCR体系(25 μL):超纯水16.5 μL,25 mg·mL-1MgCl21.5μL,10×Buffer 2.5 μL,10 mmol·L-1dNTP 2.0 μL,10 μmol·L-1PrimerⅠ 0.5 μL,10 μmol·L-1primerⅡ 0.5μL,50 ng·μL-1模板1.0 μL,5 U·μLTaq酶0.5 μL。PCR反应程序:94 ℃预变性8 min;94 ℃变性30 s,52.5 ℃退火30 s,72 ℃延伸1 min,30个循环;72 ℃延伸8 min;4 ℃终止反应。1%琼脂糖凝胶电泳检测。

图1 P2300-Pa-8e植物表达质粒结构示意图
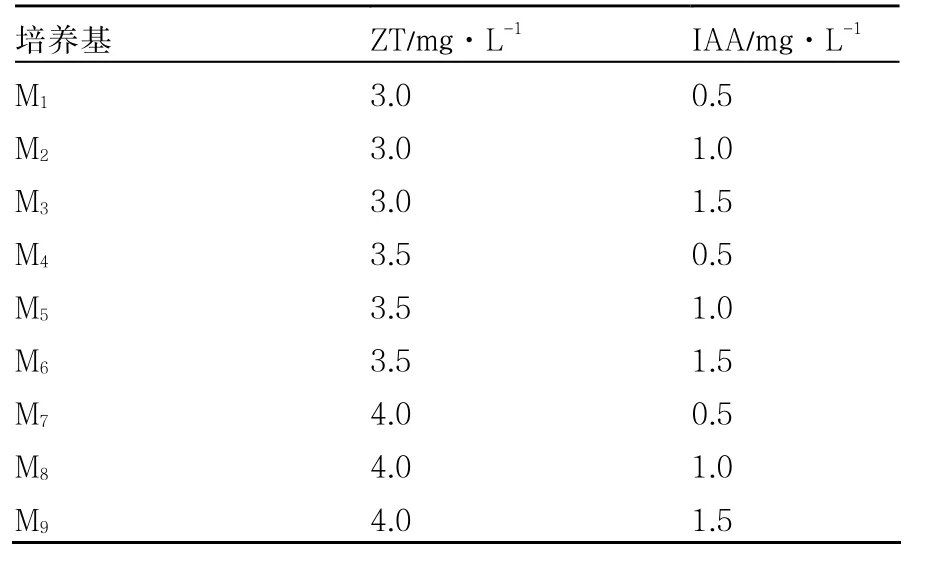
表1 不同薯块再生培养基的激素配比
1.2.5 Southern-blot杂交检测 用SacⅠ酶切转基因植株DNA,将酶切产物进行琼脂糖凝胶电泳,经变性、中和,用高盐转移法(萨姆布鲁克,2002)将胶上的样品转移到尼龙膜上,以目的基因P2300-pa-8e为探针,采用地高辛试剂盒进行杂交、洗膜、显影。
1.2.6 转基因植株的移栽 将通过Southern-blot杂交检测的转基因植株培养15~20 d,炼苗后移入小盆中,放入温室培养,7 d左右转入网室培养。
1.2.7 抗虫试验 用转基因小薯与对照小薯分别喂养暗黑鳃金龟幼虫(3~4日龄)、华北大黑鳃金龟幼虫(3~4日龄)、黄粉虫幼虫和超级大麦虫幼虫。暗黑鳃金龟(3~4日龄)、华北大黑鳃金龟(3~4日龄)的幼虫采用室内杀虫活性测定(王容燕 等,2007),黄粉虫、超级大麦虫的幼虫采用饲料混合法(张杰 等,2002)进行生物测定。每处理 24头初孵幼虫,3次重复,置于光照培养箱中培养,分别于48、72、96 h记录死虫数,并计算校正死亡率。

2 结果与分析
2.1 共培养时间的确定
共培养时间的长短对遗传转化效率有影响,而且不同物种及外植体材料与农杆菌的最佳共培养时间也不同。从图2可以看出,早大白和克新18号薯片的共培养时间均在第3天时抗性愈伤组织诱导率最高,分别为92.16%和97.77%。
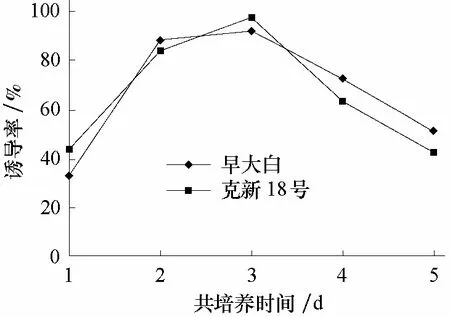
图2 共培养时间对遗传转化的影响
2.2 薯块再生培养基的确定
以MS为基本培养基,以早大白为试材,研究添加IAA和ZT两种激素对诱导马铃薯再生率的影响。试验结果表明(表2),使用M8培养基时,获得的出芽率最高,高达 93.55%;而使用M1培养基培养的薯块出芽率最低,仅有33.33%。因此,本试验条件下最优薯块再生培养基为M8(MS+4.0 mg·L-1ZT +1.0 mg·L-1IAA)。
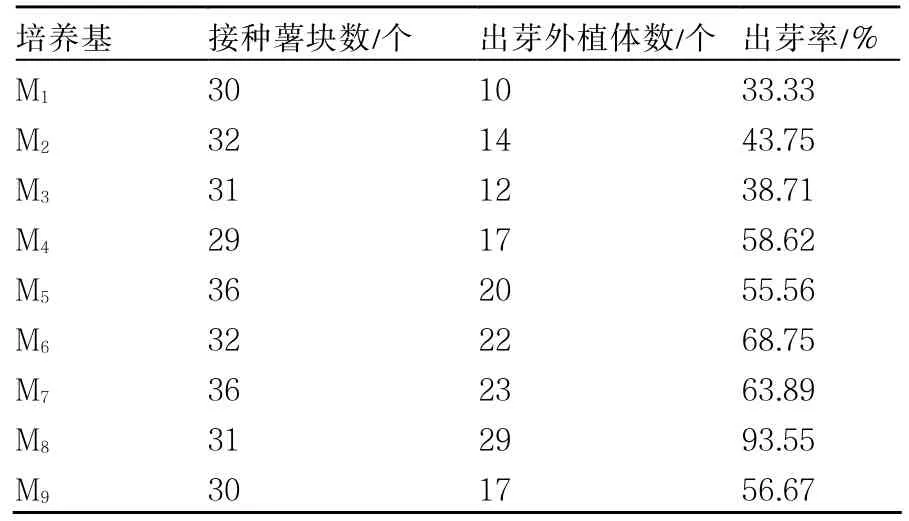
表2 不同激素浓度配比对早大白薯块生长的影响
2.3 转基因植株与薯块的获得
马铃薯薯块经培养20~30 d后分化出芽。早大白接种薯块 36块,诱导产生再生植株 42株,有19株通过生根阶段筛选,抗性植株再生频率为52.78%;克新18号接种薯块32块,诱导产生再生植株54株,有21株通过生根阶段筛选,抗性植株再生频率为65.63%。将抗性植株移入网室,9月收获薯块(图3)。

图3 农杆菌介导克新18号薯块遗传转化再生植株与小薯的获得
2.4 PCR检测
对早大白和克新 18号所获得的抗性植株进行PCR鉴定,得到了约500 bp的特异性条带(图4),与阳性对照质粒的扩增结果一致,阴性对照没有特异性扩增条带。PCR检测结果初步表明,P2300-pa-8e基因已整合到受体基因组中。
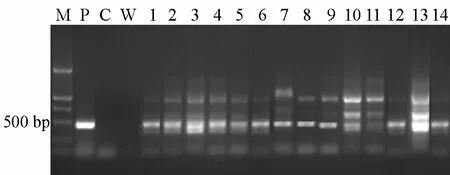
图4 抗性植株PCR检测
2.5 Southern-blot杂交检测
将PCR检测呈阳性的植株进行Southern-blot杂交,使用SacⅠ对转基因植株DNA进行酶切,在5株早大白、10株克新18号PCR阳性植株中,4株早大白、6株克新18号有杂交信号(图5),其中4株为双拷贝,6株为单拷贝。不同植株外源基因插入位点不同,说明不同植株之间是相对独立的转基因植株。
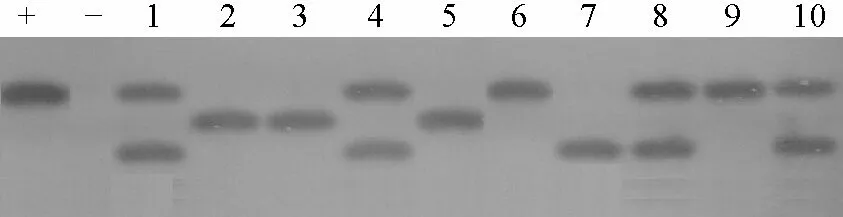
图5 转化植株Southern-blot杂交检测
2.6 抗虫试验
以非转基因马铃薯为对照,测定P2300-pa-8e基因表达产物对暗黑鳃金龟、华北大黑鳃金龟、黄粉虫、超级大麦虫幼虫的毒力。结果表明,对暗黑鳃金龟、华北大黑鳃金龟龄幼虫7 d的校正死亡率分别为26.38%、34.72%;14 d的校正死亡率分别为47.22%、55.56%;而对黄粉虫、超级大麦虫幼虫均未表现活性。
3 结论与讨论
选择马铃薯微型薯作为外植体,遗传转化过程中具有再生速度快、出芽率高、易于操作等优点。
培养基中激素的组成和浓度是影响植株再生的关键,生长素和分裂素比例合适可以提高胚状体发生频率。从诸多马铃薯再生培养的报道中可以看出,所使用的外源激素大相径庭(张宁等,2004),用一种或几种培养基并不适合所有的品种。因此,需针对不同基因型选择诱导再生的激素种类和浓度。本试验使用MS+4.0 mg·L-1ZT+1.0 mg·L-1IAA为薯块再生培养基时获得的出芽率最高,高达93.55%。
适当的共培养时间能提高转化效率,本试验结果表明,共培养时间以外植体周围刚出现白色菌落时为最佳(一般共培养2~3 d),早大白和克新18号最佳共培养时间均为3 d。这样既可提高转化频率,又易于去除农杆菌。达到最佳生长状态的共培养时间则由农杆菌菌液的生长状态、菌液浓度、外植体类型及培养条件等决定。
对诱导生根的40株马铃薯抗性植株进行PCR检测,有14株呈PCR阳性反应,但都出现了非特异性扩增条带,其原因可能是:① 引物与靶序列不完全互补或引物聚合形成二聚体。② Mg2+离子浓度过高、退火温度过低,及PCR循环次数过多。③ 酶的质和量,有些来源的酶易出现非特异条带,而另一来源的酶则不出现;酶量过多有时也会出现非特异性扩增。
经Southern-blot检测,有10株为转基因植株,表明在本试验中存在假转化体。分析原因可能有:① 外植体的表层细胞稳定表达外源选择抗性基因,为其他一些非转化细胞提供了一道屏障,逃避了选择压力的影响而继续分裂和分化。② T-DNA进入细胞后并没有整合到受体基因组中,而是以游离状态存在,导致PCR呈阳性反应。因此只有进行植物基因组的Southern-blot分析,才可以准确地了解外源基因在转化植株基因组中的整合情况。
本试验通过对薯块的抗虫试验,结果表明转基因马铃薯对鳞翅目害虫黄粉虫、超级大麦虫无活性,对鞘翅目害虫暗黑鳃金龟和华北大黑鳃金龟表现出抗性,为马铃薯抗性品种的培育提供了新的材料和方法。
丛培琳,卢翠华,邸宏,石瑛,张丽莉,姜丽丽,李文滨.2008.Bt-CryV基因对马铃薯的遗传转化.东北农业大学学报,39(9):16-20.
姜丽丽,邸宏,林忠平,胡鸢雷,吴韩英,崔少彬,姜丽静,卢翠华.2009.蓝铜蛋白类似基因BcBCP1转化马铃薯及其抗旱性的改良.中国蔬菜,(2):7-11.
卢翠华,邸宏,张丽莉.2009.马铃薯组织培养原理与技术.北京:中国农业科学出版社.
萨姆布鲁克 J,拉塞尔 D W.2002.分子克隆试验指南.3版.北京:科学出版社.
王容燕,王金耀,宋健,曹伟平,杜立新,冯书亮,宋福平,张杰.2007.铜绿丽金龟的室内人工饲养.昆虫学报,50(1):20-24.
姚庆学,张勇,丁岩.2003.金龟子防治研究的回顾与展望.东北林业大学学报,31(3):64-66.
张杰,宋福平,李长友,陈中义,檀建新,黄大昉.2002.对鞘翅目害虫高毒力Bt基因cry3Aa7的分离克隆及表达研究.中国农业科学,35(6):650-653.
张宁,司怀军,李学才,王蒂.2004.根癌农杆菌介导的马铃薯高效遗传转化体系的研究.中国马铃薯,18(3):132-135.
James C.2008.Global status of commercialized Biotech/GM crops.ISAAA Brief No.39.Ithaca NY:ISAAA
Yu H,Zhang J,Huang D F,Gao J G,Song F P.2006.Characterization ofBacillus thuringiensisstrain Bt185 toxic to the Asian cockchafer:Holotrichia parallela.Curr Microbiol,53(1):13-17.
猜你喜欢
——粪金龟
科学大众(2022年19期)2022-10-30
文史博览(2020年11期)2020-12-01
四川动物(2020年3期)2020-06-11
吉林林业科技(2020年2期)2020-04-15
天然产物研究与开发(2018年4期)2018-05-07
农村百事通(2018年21期)2018-01-03
中国组织化学与细胞化学杂志(2017年1期)2017-06-15
中国农业文摘-农业工程(2017年2期)2017-03-31
今日农业(2017年7期)2017-02-02
河南农业(2017年1期)2017-02-01
