
青钱柳营养器官的解剖结构及其生态适应性1)
2011-05-31 08:58洑香香
东北林业大学学报 2011年10期
王 玉 赵 虎 洑香香
(南京林业大学,南京,210037)
青钱柳[Cyclocarya paliurus(Batal.)Iljinskaj]系双子叶植物胡桃科(Juglandaceae)青钱柳属高大乔木,又名青钱李(江西)、山麻柳(四川)、甜茶树(贵州)、一串钱(湖北),是我国特有的单种属植物,国家重点保护的三类濒危植物之一。它广泛分布于安徽、江苏、浙江、江西、福建、台湾、湖北、四川、贵州等地的海拔420~2 500 m的山区、溪谷或石灰岩山地[1]。由于青钱柳主要分布在中国,因此国外涉及青钱柳的研究比较少,仅限于对青钱柳药用价值的研究[2]。国内对青钱柳的研究较多,但主要集中在种子休眠[3-4]、组织培养[5]、扦插繁殖[6]、次生代谢物质的提取和鉴定[7]等方面,而对营养器官解剖结构的研究仅有叶的相关报道[8]。笔者利用扫描电镜和常规石蜡切片技术对青钱柳的根、茎、叶进行了系统的观察和研究,并试图探讨营养器官的解剖结构与生态适应性的关系,为青钱柳的推广栽培提供科学依据。
1 材料与方法
成熟叶片和2年生枝采自南京林业大学校园内栽培的6年生青钱柳植株,当年生枝和根采自温室栽培的当年生青钱柳植株。
扫描电镜制片:将叶片中部切成1 cm×1 cm小块,经70%FAA固定液固定24 h,系列酒精脱水,用双面胶带贴在盖玻片上,置于ALPHA1-2 Ldplus型冷冻干燥机干燥3.5 h,渡膜喷金后,在扫描电镜(quanta-200)下观察并照相。
石蜡制片:将叶片切成4 mm×6 mm的小块,根、茎截取5 mm长的小段,切好的材料用清水冲洗后投入FAA固定液中固定。固定24 h后,将根和茎转入软化剂中进行软化,2年生枝用10%乙二胺软化(软化2个月),当年生枝和根用50%甘油+50%乙醇(质量分数95%)软化(主根软化2个月,侧根和当年生枝软化2周)。常规石蜡切片法切片,切片厚度为8~12 μm,番红―固绿双重染色,中性树胶封片,OlympusCX-41型光学显微镜观察并拍照。
2 结果与分析
2.1 叶的形态与解剖结构
扫描电镜下叶的表面形态:扫描电镜下,青钱柳叶表皮细胞外壁的垂周壁呈波状(图1a、图1b)。上、下表皮均具有非腺毛和腺毛,下表皮分布的较上表皮多,且叶脉处的非腺毛和腺毛较其他地方多。气孔仅分布于叶下表皮,略外凸,长15~20 μm,宽8~10 μm,气孔密度约为 140 个/mm2,气孔和表皮毛基部周围的角质层纹饰呈辐射状(图1a、图1b)。非腺毛单列,长50~100 μm。腺毛具有一个短的柄细胞和多个膨大的头细胞,下陷(图1a、图1b)。
叶的解剖结构:青钱柳叶为异面叶。从横切面上看,厚290~330 μm,由表皮、叶肉和叶脉组成。表皮分为上表皮和下表皮,仅由一层细胞组成,细胞长方形,排列紧密,无胞间隙,厚约40 μm(图1c、图1d)。栅栏组织与海绵组织分化明显,栅栏组织位于上表皮下方,由一层细胞构成,细胞长柱状,排列紧密整齐;栅栏组织中分布着簇晶,体积较大,数量较多(图1c)。海绵组织由2~3层细胞组成,细胞大小不等,细胞间隙大,排列疏松,有少量的簇晶分布(图1d)。叶肉组织内有小的维管束,束鞘纤维木化,成环(图1d)。主脉中维管束环状,木质部在近轴面,较为发达,韧皮部在远轴面(图1e、图1f),木质部与韧皮部之间有微弱的的形成层,韧皮部的外围分布有少量簇晶(图1f、图1g)。值得注意的是,在正常维管束的上方还具有一个副维管束(图1g)。
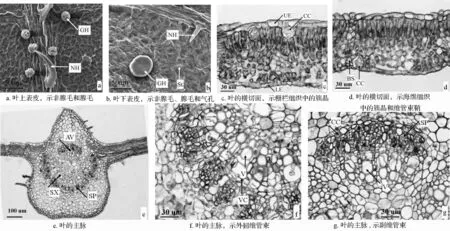
图1 叶的形态与解剖结构
2.2 茎的解剖结构
青钱柳幼茎由表皮、皮层与维管柱构成。表皮细胞1层,横切片观察表皮细胞呈长方形,排列紧密整齐,没有细胞间隙,外突形成腺毛(图2a)。皮层主要由薄壁细胞组成,皮层外部紧贴表皮的一层细胞开始活动,形成木栓形成层,皮层内部薄壁细胞排列紧密,细胞之间分布着少量簇晶(图2b)。维管柱由维管束、髓和髓射线组成。维管束为外韧维管束,在茎中排列为一轮,5~6个。韧皮部主要由筛管、伴胞、韧皮纤维和薄壁细胞组成,木质部主要由导管、管胞、木纤维和木薄壁细胞组成,导管呈辐射状分布在薄壁细胞中,3~5个导管形成一列。形成层位于韧皮部和木质部之间,由2~3层较小的长方形细胞构成,彼此排列紧密,形成层周围也有簇晶存在。髓处于茎中央,由薄壁细胞构成,所占比例最大,位于中央的细胞较大,近维管束处的细胞较小,无细胞间隙(图2a)。
青钱柳老茎由周皮和次生维管组织构成。周皮呈褐色,由木栓层、木栓形成层和栓内层组成。木栓形成层起源于紧贴表皮的一层皮层细胞,以平周分裂为主,向内外分别产生栓内层和木栓层(图2b)。木栓层排列整齐,由3~4层细胞构成,细胞呈长方形,有栓质化增厚(图2c)。次生维管组织包括次生韧皮部、维管形成层和次生木质部,次生韧皮部含有大量的簇晶和纤维(图2d),次生木质部由导管、木纤维和少量的木薄壁细胞组成,木纤维数量较多,成径向规则排列,导管孔径较大,数量较多,呈径向排列(图2e)。髓的一部分细胞死亡破坏,而另一部分细胞未被破坏,形成片状髓(图2f)。
2.3 根的解剖结构
根的初生结构:青钱柳根的初生结构由表皮、皮层和维管柱组成(图3a)。表皮由一层排列紧密的表皮细胞组成,细胞呈砖型,细胞壁薄,不具气孔。皮层位于表皮之内,在横切面上占相当大的部分,由多层薄壁细胞组成,有胞间隙,含有少量的簇晶(图3b);外皮层位于皮层的最外层,即紧贴表皮的一层,细胞排列紧密;内皮层位于皮层的最内层,有一层排列紧密的细胞组成,比皮层细胞小,无胞间隙,具有凯氏带(图3c)。维管柱位于根的中心,内皮层以内的部分,在横切面上占有较小的面积,包括中柱鞘和初生维管组织。中柱鞘是维管柱最外面的细胞层,紧贴着内皮层,由一层排列紧密的薄壁细胞组成;初生维管组织位于根的中心,包括初生木质部和初生韧皮部,二者相间排列,各自成束,中间有薄壁组织相隔,初生木质部为四原型,位于维管柱的中央,呈星状排列(图3c)。
根的次生结构:青钱柳成熟主根从外到内由周皮和次生维管组织组成(图3d)。周皮较厚,由木栓层、木栓形成层和栓内层组成。最外层为4~5层木栓层细胞,细胞为规则的长方形,排列整齐紧密,有些木栓层细胞形状已经被挤毁;木栓形成层不明显,其内为1~2层栓内层细胞。次生维管组织包括次生韧皮部、维管形成层和次生木质部。次生韧皮部主要由韧皮薄壁细胞组成,细胞形状不规则,呈近方形,排列紧密,含有大量的簇晶和纤维(图3e)。维管形成层由3~5层细胞构成,细胞呈扁长形(图3d)。在次生木质部中,导管口径大小不一,星散分布;维管射线明显,多由1列细胞构成,少数为2列细胞;木薄壁细胞排列整齐紧密,含有少量簇晶(图3f)。
3 讨论
青钱柳的自然分布表现出广泛的适应性,南及广东,北达陕西,东到台湾,西至云南,多生于海拔420~2 500 m山区、溪谷、林缘、林内或石灰岩山地,喜生于温暖、湿润肥沃、排水良好的酸性红壤、黄红壤之上,适生于湿度较大的环境中[9],是一种耐水湿树种。在对青钱柳天然林分调查时发现,其干形通直,树体高大,为上层林木的优势树种,成年树表现出明显的阳性树种特性;而林中自然更新的幼苗常处于土层较厚、透光率低的大树林下,幼苗表现出极强的耐荫性;而幼树则出现 在有一定光照的林缘、林中间隙处。

图2 茎的解剖结构
3.1 营养器官的解剖结构与湿生环境相适应的特征
生长于不同生态环境中的植物,常表现出不同的形态结构,这通常被认为是植物对特殊生境的演化适应[10]。
叶是植物进行同化和蒸腾作用的主要器官,与周围环境有着密切的关系。在长期外界环境的影响下,叶在形态结构上的变异性和可塑性最大,即叶对生态条件的反映最为明显[11]。青钱柳的叶片与多数双子叶植物的叶片相类似,为典型的异面叶。疏松的海绵组织是对中生、湿生环境的适应[12];主脉维管束为外韧型,在正常维管束的上方还具有一个副维管束,如此发达的叶脉大大提高了对水分的运输效率,这些特点是与其沟谷中的生境相适应的[13]。
在土壤渍水的情况下,植物根系所需的氧气主要通过茎叶的胞间隙系统向根系运输,所以茎叶胞间隙的大小与输氧量的多少密切相关[14]。茎中大的髓腔可以作为通气组织,在一定范围内提高它的耐渍性[15]。青钱柳茎次生结构的中央形成片状髓,这种结构也可以作为通气组织,使青钱柳能适应一定程度的水淹。
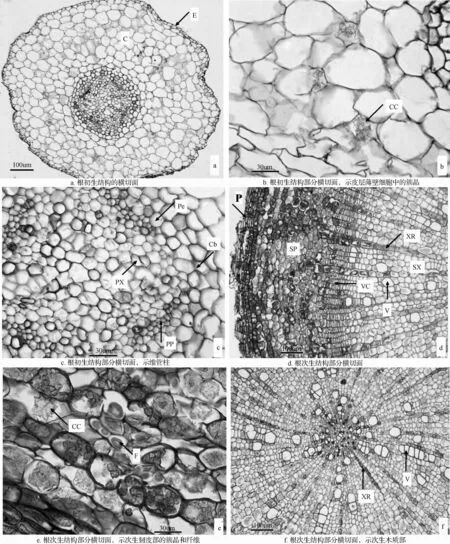
图3 根的解剖结构
3.2 营养器官的解剖结构与阳生及阴生环境相适应的特征
3.2.1 成年树与阳生环境相适应的特征
青钱柳成年树喜光,为阳性树种,其叶的解剖结构具有以下特点:①角质层是陆生植物体表抵御外界环境不良影响的屏障,青钱柳叶角质层较厚[8],可以反射和适应强烈阳光的照射,避免高海拔下强烈阳光灼伤叶片,说明青钱柳可适应不同海拔高度光照强度和光质的剧烈变化。②青钱柳叶的上、下表皮均具有非腺毛和腺毛,研究表明,表皮附属物能够反射阳光,减少叶片表面空气的流动,降低蒸腾作用,防止水分过度丧失[16],有利于维持植物正常的生理代谢。③栅栏组织细胞长柱形,排列紧密,有利于提高光能利用率[13],这与其喜光性相适应。
3.2.2 幼苗与阴生环境相适应的特征
青钱柳叶片气孔分布于下表皮,且外凸,这一特征可以反映出其叶是适应于潮湿阴生环境的。气孔外凸现象可使其开口增大,能够减小因外被附属物所引起的气孔阻力,从而提高叶片与外界环境的气体交换能力,增强叶片对二氧化碳的摄入,以提高光合作用速率[17],是植物对林下低二氧化碳和低光照的适应。
青钱柳茎和根的初生结构中机械组织和输导组织都不够发达。这一特征使幼苗不能在高温强日照环境下生存,只能在阴湿条件下叶片才能得到充足的水分,满足光合作用和蒸腾作用的需要[18]。
综上所述,青钱柳营养器官的解剖结构一方面表现出湿生植物的结构特点,另一方面又具有一些阳生和阴生植物的结构特点,反映出植物结构与环境的统一性。因此在引种栽培青钱柳时最好选择有沟谷的迎风向阳的山坡上育苗,在相对湿度大的沟谷两旁、山坡中下部生长最为良好,在高温强日照天气下,幼苗可适当遮荫,成年树喜光,易在疏林中生长。
[1]中国科学院中国植物志编辑委员会.中国植物志:21卷[M].北京:科学出版社,1979.
[2]Kurihara H,Fukami H,Kusumoto A,et al.Hypoglycemic action of Cyclocarya paliurus(Batal.)Iljinskaj in normal and diabetic mice[J].Bioscience Biotechnology and Biochemistry,2003,67(4):877-880.
[3]史晓华,徐本美,黎念林,等.青钱柳种子休眠与萌发的研究[J].种子,2002(5):5-7.
[4]尚旭岚,徐锡增,方升佐.青钱柳种子次生休眠的发生及贮藏物质的变化[J].南京林业大学学报:自然科学版,2006,30(2):99-102.
[5]胡冬南,上官新晨,刘亮英,等.青钱柳茎段离体培养研究[J].湖北农业科学,2009,48(6):1300-1303.
[6]谢风,潘斌林,胡冬南,等.青钱柳嫩枝扦插影响因素的研究[J].安徽农业科学,2009,37(9):4014-4015.
[7]李俊,黄锡山,陆园园,等.青钱柳化学成分的研究[J].中成药,2008,30(2):238-240.
[8]孙同兴,徐丽丽.青钱柳叶的结构鉴定[J].中草药,2006,37(2):271-273.
[9]连雷龙.青钱柳的栽培技术[J].林业科技开发,2003,17(3):51-52.
[10]伊稍K.种子植物解剖学[M].2版.李正理,译.上海:上海科学技术出社,1982:245-249.
[11]王怡.三种抗旱植物叶片解剖结构的对比观察[J].四川林业科技,2003,24(1):64-67.
[12]Lee D W,Bone R E,Tersis S.Corredates of leaf optical properties in tropical forest sun and extreme-shade plants[J].American Journal of Botany,1990,77:370-380.
[13]杨开军,张小平,张兴旺,等.稀有植物香果树叶解剖结构的研究[J].植物研究,2007,27(2):195-198.
[14]侯小改.芝麻不同抗性品种营养器官结构的比较研究[J].河南农业大学学报,1994,28(4):399-403.
[15]王晖,周守标,史国芹.假俭草和结缕草营养器官结构对抗逆性的影响[J].植物研究,2007,27(6):701-707.
[16]贺金生,陈伟烈,王勋陵.高山栎叶的形态结构及其与生态环境的关系[J].植物生态学报,1994,18(3):2l9-227.
[17]何涛,吴学明,贾敬.青藏高原高山植物的形态和解剖结构及其对环境的适应性研究进展[J].生态学报,2007,27(6):2574-2583.
[18]袁王俊,张维瑞,尚富德.黄连营养器官解剖结构与其阴生环境相关性研究[J].河南大学学报:自然科学版,2007,37(2):184-186.
猜你喜欢
林业科学(2020年10期)2020-11-30
西藏人文地理(2020年4期)2020-11-19
农业工程技术·综合版(2020年1期)2020-04-23
蔬菜(2018年12期)2018-01-16
浙江大学学报(农业与生命科学版)(2017年1期)2017-04-17
江苏农业科学(2017年1期)2017-02-27
天津农业科学(2015年11期)2015-12-03
湖北农业科学(2014年16期)2014-10-20
江苏农业科学(2014年7期)2014-09-02
山东农业科学(2014年4期)2014-07-18
