
绒毛状烟草基因组中SSR位点的信息分析
2011-05-25 06:52王卫锋晁江涛龚达平李凤霞陈雅琼孙玉合
中国烟草科学 2011年3期
王卫锋,晁江涛,龚达平,王 鲁,李凤霞,陈雅琼,2,孙玉合*
(1.农业部烟草类作物质量控制重点开放实验室,中国农业科学院烟草研究所,青岛 266101;2.中国农业科学院研究生院,北京 100081)
简单重复序列( simple sequence repeats,SSR)又称微卫星DNA、短串联重复序列,一般为以 1~6个碱基为核心序列,首尾相连组成的串联重复序列[1]。SSR位点广泛存在于真核生物基因组中,不同遗传材料在重复次数上的差异可导致 SSR位点长度的多态性,在SSR位点两侧有一段相当保守的单拷贝序列,根据此保守序列设计PCR引物能够精确检测出SSR位点的长度多态性。以SSR位点的长度多态性为基础,利用PCR和电泳技术开发出的SSR分子标记具有对基因组覆盖度好、数量丰富、多态性高、共显性标记、实验简单以及重复性好等特点[2]。
SSR分子标记已在植物遗传分析中得到广泛应用,如遗传作图、分子标记辅助选择、群体遗传分析以及分类学等方面,在花生、玉米、大麦[3-5]等作物中利用SSR标记已构建了遗传连锁图谱;Korzun等[6]利用SSR标记对小麦矮杆基因Rht8进行了定位;Prevost等[7]对34个马铃薯品种构建了SSR指纹图谱;SSR分子标记在小麦、大麦等作物的遗传多样性分析中也得到了成功应用[8-9]。另外,在烟草中也利用SSR标记开展了相关研究,如范静苑等[10]成功运用SSR分子标记分析了烟草CMV抗性基因的遗传连锁距离;Moon等[11]运用SSR分子标记分析了54份烟草属材料的遗传多样性。
烟草分子标记研究多以RAPD和AFLP为主,主要应用于野生种质的抗病基因鉴定[12-18],限于这些分子标记多态性较低,用其构建烟草遗传图谱受到了很大限制[19-21]。对烟草基因组的测序极大地促进了烟草 SSR分子标记的开发,Bindler等[22]从Tobacco Genome Initiative(TGI)公布的烟草基因组序列信息中搜索到282个SSR分子标记,并利用这些标记构建了第一张烟草的遗传连锁图。在本研究中,通过软件搜索绒毛状烟草基因组序列,筛选到一批SSR位点,同时对其组成、分布及特征进行了分析,以期为开发新的烟草SSR分子标记提供理论依据。
1 材料与方法
利用软件SSRFINDER(http://www.maizemap.org/bioinformatics/SSRFINDER/)搜索绒毛状烟草基因组DNA的序列数据,条件为:重复单元长度2~5 bp,重复次数为10~60。
2 结 果
2.1 SSR位点的数量与分布
利用软件 SSRFINDER对绒毛状烟草基因组DNA的序列数据进行搜索,共找到符合条件的SSR位点6441个,涉及5227个Scaffold,SSR位点的序列总长303963 bp(表1)。从SSR位点的数量上看,三碱基重复基元的数量最多,占49.6%;其次是二碱基重复基元,占45.2%;四碱基重复基元和五碱基重复基元数量很少,分别只占2.8%和2.4%(图1)。二碱基重复基元、三碱基重复基元、四碱基重复基元和五碱基重复基元的 SSR位点的碱基总长分别是112678 bp、167889 bp、11616 bp和11780 bp,位点的平均长度分别是39 bp、53 bp、66 bp和75 bp。
从重复次数看,10~20重复的位点有4809个,占74.6%;21~30重复的位点有963个,占15.0%;31~40重复的位点有349个,占5.4%;41~50重复的位点有205个,占3.2%;51~60重复的位点有113个,占1.8%(图2)。
2.2 重复单元的类型、数量及特征
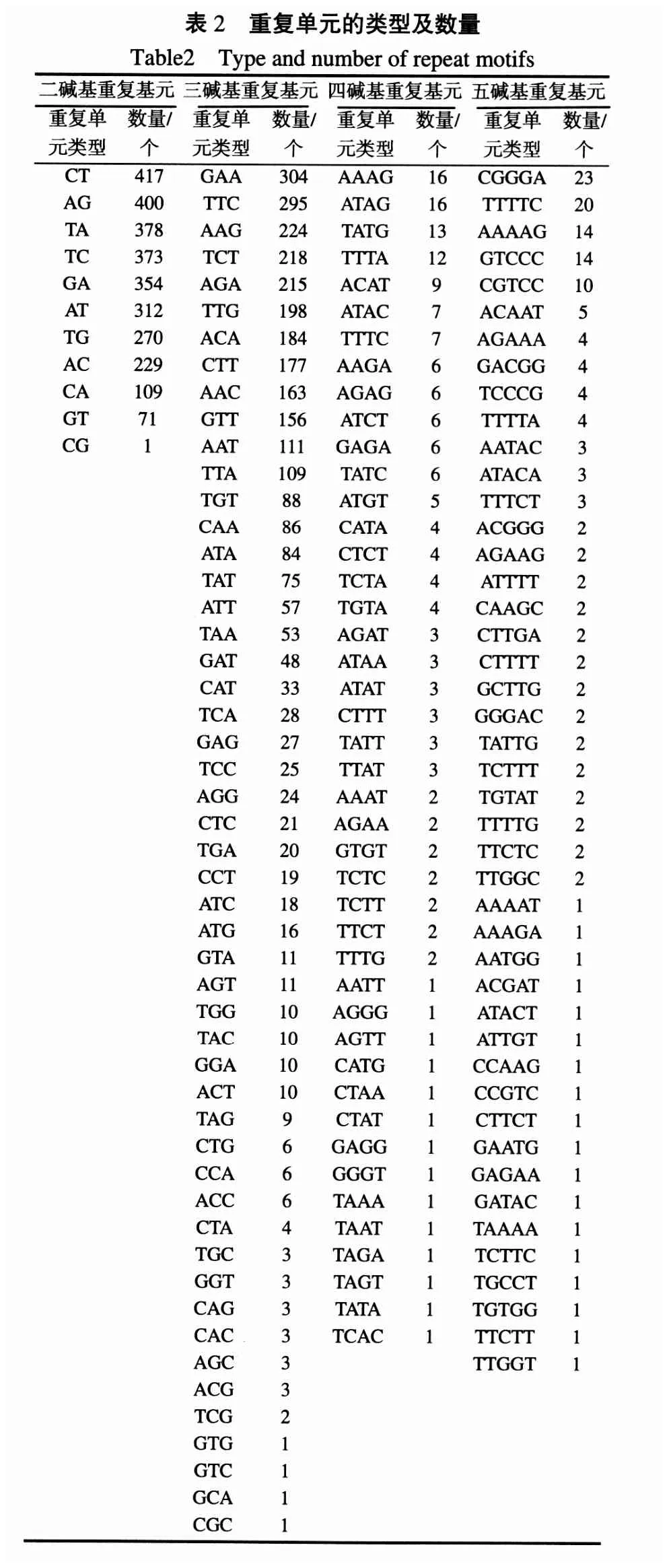
在6441个SSR位点中,二碱基重复基元单元共出现11种类型,分别是CT(417个)、AG(400个)、TA(378个)、TC(373个)、GA(354个)、AT(312个)、TG(270个)、AC(229个)、CA(109个)、GT(71个)、CG(1个);三碱基重复基元单元共出现 51种类型,数目最多的前 10种依次是GAA(304个)、TTC(295个)、AAG(224个)、TCT(218个)、AGA(215个)、TTG(198个)、ACA(184个)、CTT(177个)、AAC(163个)、GTT(156个);四碱基重复基元单元共出现44种,数目最多的前10种依次是AAAG(16个)、ATAG(16个)、TATG(13个)、TTTA(12个)、ACAT(9个)、ATAC(7个)、TTTC(7个)、AAGA(6个)、AGAG(6个)、ATCT(6个);五碱基重复基元单元共出现45种类型,数目最多的前10种依次是 CGGGA(23个)、TTTTC(20个)、AAAAG(14个)、GTCCC(14个)、CGTCC(10个)、ACAAT(5个)、AGAAA(4个)、GACGG(4个)、TCCCG(4个)、TTTTA(4个)(表2)。A、T、C、G在所有SSR位点中所占的比例分别为33.1%、33.4%、16.6%和16.9%(图3),GC含量为33.5%。
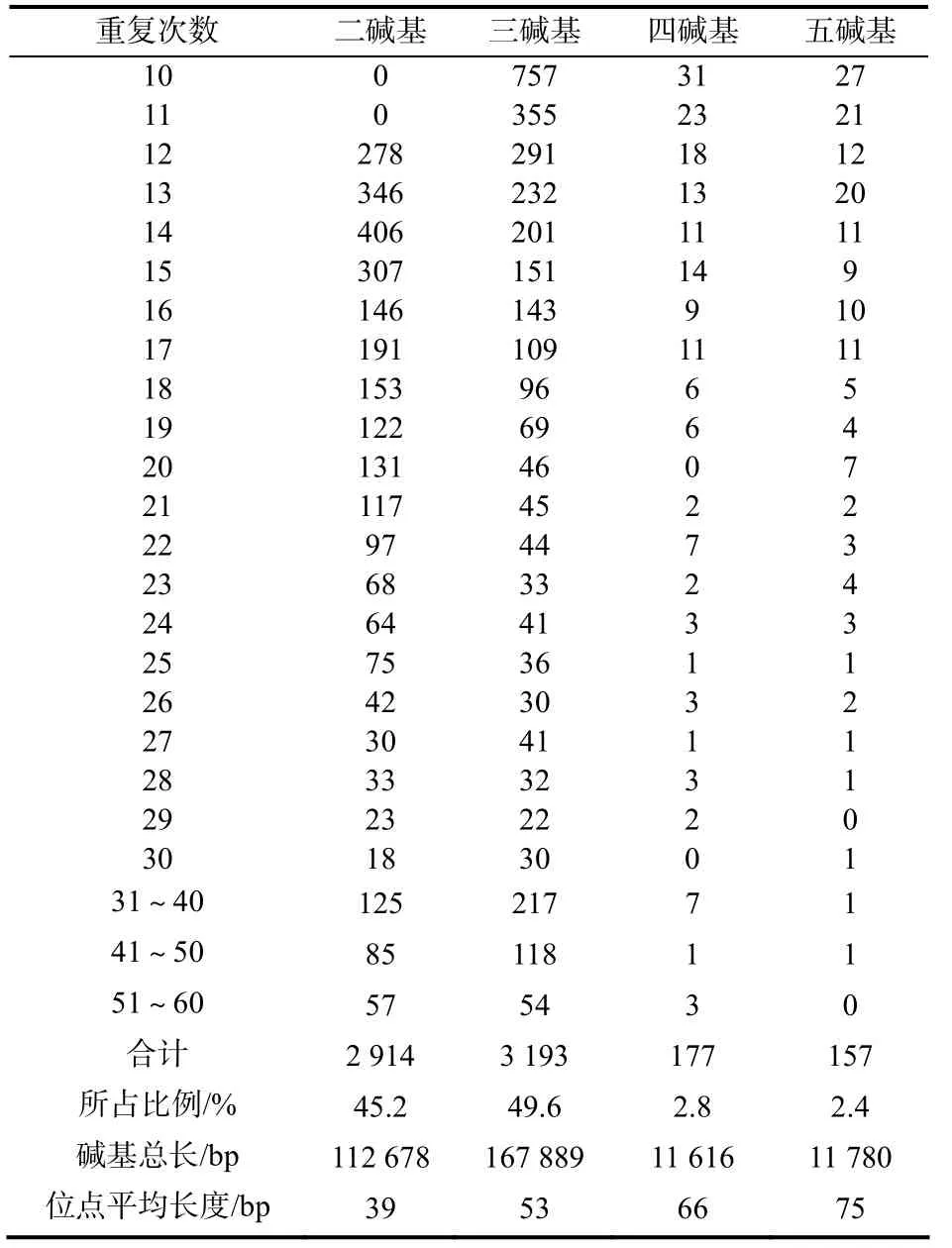
表1 绒毛状烟草基因组SSR位点的数量与分布Table1 Distribution of SSR loci in N.tomentosiformis Goodspeed genome DNA
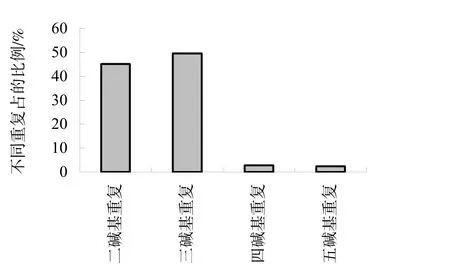
图1 4种重复单元的分布比例图Fig.1 Distribution of four repeat motifs
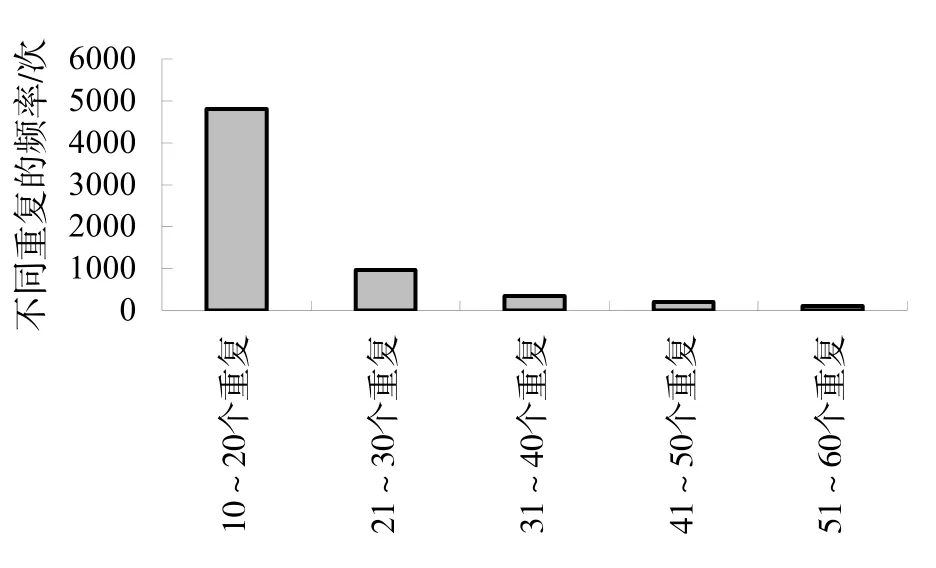
图2 重复次数分布图Fig.2 Distribution of the number of repeats
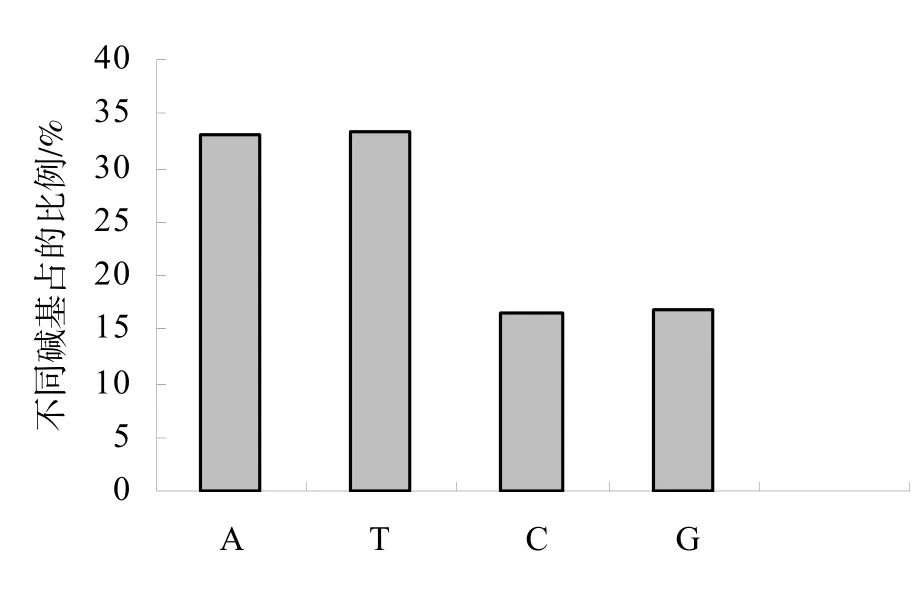
图3 SSR位点中四种碱基的比例Fig.3 Distribution of A, T,C and G in SSR loci
3 讨 论
根据测序结果,绒毛状烟草基因组大小为2.6Gb(待发表),本研究从中筛选 SSR总长约为300 kb,约占基因组的万分之1.2,换句话说绒毛状烟草基因组平均每400 kb出现一个SSR。而本研究并未计算单碱基重复基元和六碱基重复基元的SSR,所以SSR实际出现的频率还会略高一些。不同物种间,SSR的数量、重复次数、重复单元以及在染色体上的分布均存在很大的差异[9]。在植物中,SSR的出现频率与基因组大小、重复DNA所占比例呈负相关,而与转录区所占的比例、低拷贝序列的出现频率成显著正相关[23]。
?
绒毛状烟草基因组的GC含量为38.9%,其中SSR序列的GC含量为33.5%,明显低于全基因组的GC含量。SSR序列显示出显著的碱基偏好性,A/T的频率显著高于G/C。二、三碱基重复基元的SSR共6107个,占SSR总体的94.8%。而(CG)n、(GTG)n、(GTC)n、(GCA)n、(CGC)n分别只出现了1次,说明这些重复单元的SSR在绒毛状烟草基因组中非常少见。从总体来看,各种重复类型的 SSR位点都存在着长度越长出现的频次越少的趋势。目前,对于植物SSR序列信息的分析,大多来源于EST数据[24-30],由于EST-SSR来自于转录区域,一般保守性较高,通用性好。Cho[31]等人对水稻的研究结果表明,来源于基因组的SSR比来源于EST的SSR具有更丰富的多态性。研究发现,当SSR长度大于等于20 bp时,不同品种间显示出较高的多态性[32],而且高级基元的SSR多态性比低级基元的低[33]。在这6441个SSR位点中,二、三、四、五重复基元SSR位点的平均长度分别为39 bp、53 bp、66 bp和75 bp,其中二、三碱基重复基元的总和占总体的94.8%,表明这些SSR具有较高的利用价值。用PRIMER5.0对这6441个SSR设计引物,其中6020可获得满足条件的引物。至于哪些引物可用于开发新的烟草 SSR分子标记还有待进一步研究。
[1]Beckmann J S, Soller M.Toward a unified approach to genetic mapping of eukaryotes based on sequence tagged microsatellite sites[J].Bio Technology, 1990, 8: 930-932.
[2]Powell W, Machray G C, Provan J.Polymorphism revealed by simple repeats[J].Trends Plan Sci, 1996, 1:215-222.
[3]洪彦彬,梁炫强,陈小平,等.花生栽培种 SSR遗传图谱的构建[J].作物学报,2009,35(3):395-402.
[4]向道权,曹海河,曹永国,等.玉米 SSR遗传图谱的构建及产量性状基因定位[J].遗传学报,2001,28(8):778-784.
[5]Thiel T, Michalek W, Varshney R K, et al.Exploiting EST database for the development and characterization of gene-derived SSR-markers in barley (Hordeum vulgareL.)[J].Theor Appl Genet, 2003, 106 (3): 11-22.
[6]Korzun V, Roder M, Wendehake K, et al.Integration of dinucleotide microsatellites from hexaploid bread wheat into a genetic linkage map of durum wheat[J].Theor Appl Genet, 1999, 98 (1): 1202-1207.
[7]Prevost A, Wilkinson M.A new system of comparing PCR primers applied to SSR fingerprinting of potato cultivars[J].Theor Appl Genet, 1998, 98 (1): 107-112.
[8]Prasad M, Varshney R, Roy J, et al.The use of microsatellites for detecting DNA polymorphism,genotype identification and genetic diversity in wheat[J].Theor Appl Genet, 2000, 100 (4): 584-592.
[9]朱宏波,方宣钧,杨仁崔.利用水稻基因组序列数据开发SSR标记的方法[J].分子植物育种,2003,1(2):273-276.
[10]范静苑,王元英,蒋彩虹,等.烟草CMA抗性鉴定及抗性基因的SSR标记研究[J].分子植物育种,2009,7(2):355-359.
[11]H S Moon, J S Nicholson, R S Lewis.Use of transferable Nicotiana tabacum L.microsatellite markers for investigating genetic diversity in the genus Nicotiana[J].Genome, 2008, 51: 547-559.
[12]Bai D, Reeleder, R, Brandle, J E.Identification of two RAPD markers tightly linked with the Nicotiana debneyi gene for resistance to black root rot of tobacco[J].Theor Appl Genet, 1995, 91: 1184-1189.
[13]Yi Y H, Ruffy R C, Wernsman E A.Identification of RAPD markers linked the wildfire resistance gene of tobacco using bulked segrement analysis[J].Tob Sci,1998, 42: 52-57.
[14]Johnson E S, Wolff M F, Wernsman E A.Marker assisted selection for resistance to black shank disease in tobacco[J].Plant Dis, 2002.86: 1303-1309.
[15]Lewis R S, Milla S R, Kernodle S P.Analysis of an introgressed Nicotiana tomentosa genomic region affected leaf number and correlated traits in Nicotiana tabacum[J].Theor Appl Genet, 2007, 114: 841-854.
[16]Milla, S R., Levin, J.S., Lewis, R.S., and Rufty, R.C.RAPD and SCAR markers linked to an introgressed gene conditioning resistance to Peronospora tabacina D.B.Adam.in tobacco[J].Crop Sci, 2005, 45: 2346-2354.
[17]Lewis R S.Transfer of resistance to potato virus Y(PVY)from Nicotiana africa to Nicotiana tabacum:possible influence of tissue culture on the rate of introgression[J].Theor Appl Genet, 2005.110: 678-687.
[18]Lewis R S , Milla S R, Levin J S.Molecular and genetic characterization of Nicotiana glutinosa L.chromosome segments in tobacco mosaic virus-resistant tobacco accessions[J].Crop Sci, 2005, 45: 2355-2362.
[19]Moon H S, Nicholson J S.Identification of AFLP markers linked to Tomato Spotted Wilt Virus resistance in tobacco[J].Crop Sci, 2007, 47: 1887-1894.
[20]Ren N, Timko M P.AFLP analysis of genetic polymorphism and evolutionary relationships among cultivated and wild Nicotiana species[J].Genome, 2001,44: 559-571.
[21]Rossi L, Bindler G, Pijnenburg H, et al.Potential of molecular marker analysis for variety identification in processed tobacco[J].Plant Var.Seed, 2001, 14: 89-101.
[22]Bindler G, Rutger van der Hoeven,el at.A microsatellite marker based linkage map of tobacco[J].Theor Appl Genet, 2007, 114: 341-349.
[23]Morgante M, Hanafey M, Powell W.Microsatellites are preferentially associated with nonrepetitive DNA in plant genomes [J].NatureGenetics, 2002, 30: 194-200.
[24]Cardle L, Ramsay L, Milbourne D, et al.Computational and experimental characterization of physically clustered simple sequence repeats in plants [J].Genetics, 2000,156:847-854.
[25]Kantety R V, Rota M L, Matthews D E, et al.Data mining for simple sequence repeats in expressed sequences tags from barley,maize,rice,sorghum and wheat[J].Plant Mol Biol, 2002, 48: 501-510.
[26]Thiel T, Michalek W, Varshney R K, et al.Exploiting EST database for the development and characterization of genderived SSR-markers in barley (Hordeum vulgare L.)[J].Theor Appl Genet, 2003, 106: 411-422.
[27]忻雅,崔海瑞,卢美贞,等.白菜 EST-SSR信息分析与标记的建立[J].园艺学报,2006,33(3):49-554.
[28]林范学,程水明,李安政,等.香菇基因组中EST-SSR的构成和分布[J].微生物学通报,2007,34(3):438-442.
[29]金基强,崔海瑞,陈文岳,等.茶树 EST-SSR的信息分析与标记建立[J].茶叶科学,2006,26(1):17-23.
[30]Jiang Dong, Zhong Guangyan, Hong Qibing.Analysis of Microsatellites in Citrus Unigenes[J].Theor Appl Genet,2003, 106: 411-422.
[31]Cho Y G, Ishii T, Temnykh S, et al.Diversity of microsatellites derived from genomic libraries and GenBank sequences in rice (Oryza sativa L.) [J].Theor Appl Genet, 2000, 100: 713-722.
[32]Temnykh S, DeClerck G, Lukashova A, et al.Computational and experimental analysis of microsattellites in rice (Oryza sativa L.)[J].Genome Res,2001, 11: 1441-1452.
[33]Dreisigacker S, Zhang P, Warburton M L, et al.SSR and pedigree analysis of genetic diversity among CIM-MYT wheat lines targeted to different mega environments [J].Crop Science, 2004, 44: 381-388.
猜你喜欢
计算机工程与应用(2023年1期)2023-01-13
世界科学技术-中医药现代化(2022年3期)2022-08-22
昆明医科大学学报(2021年3期)2021-07-22
兵工学报(2021年4期)2021-06-19
教学考试(高考生物)(2020年6期)2020-11-23
食品与生物技术学报(2020年8期)2020-01-06
学苑创造·B版(2019年5期)2019-06-14
科学24小时(2019年5期)2019-06-11
科学导报(2018年30期)2018-05-14
中国测试(2015年11期)2015-12-17
