
2株鸽源新城疫病毒的毒力鉴定和基因型分析
2011-05-18 08:40高以明张敬友王水明柯家法李超美朱雪良严继宁黄立业任晓进吴艳涛刘文博
中国动物检疫 2011年7期
高以明 ,张敬友 ,王水明 ,柯家法 ,李超美 ,朱雪良 ,张 扬 ,严继宁 ,黄立业 ,任晓进 ,吴艳涛 ,刘文博
(1.南京农业大学,江苏 南京 210095;2.南京出入境检验检疫局,江苏 南京 210001;3.江苏出入境检验检疫局,江苏 南京 210001;4.扬州大学农业部畜禽传染病学重点开放实验室,江苏扬州 225009)
鸽新城疫又称鸽Ⅰ型副黏病毒病,是由新城疫病毒(又称鸽副黏病毒Ⅰ型)引起的一种急性高度接触性败血性传染病,以肠炎、严重腹泻和神经症状为特征[1-3],其发病率为50%~100%,病死率达20%~95%。该病于1977年发生在中东地区,次年在埃及和伊拉克发生[1]。随后迅速传播到欧洲、美国、加拿大等地,1985年传播到亚洲[4-6]。1986年前后我国的鸽群中也发现了该病[7],现全国各省市均有报道[8-13]。
早期对鸽新城疫病毒的研究发现,鸽源新城疫病毒与当时鸡源新城疫病毒抗原性和生物学特性有一定的差异[1,14-15],基因型主要是Ⅵb 亚型[6,16-17],对鸡的致病性与鸡源新城疫病毒不同[3,5,8,21],然而,随着时间的推移、禽类养殖方式和规模等的改变,鸽源新城疫病毒也出现了一定的变化[18-23]。本实验对从临诊病例中分离到的两株鸽源新城疫病毒进行了毒力测定和基因型分析,现报告如下。
1 材料与方法
1.1 病毒和其他生物材料 两个鸽源NDV分离株是按照国际动物卫生组织(OIE)推荐的方法[24]在2008年从临床鸽病例中分离,命名JS-1-08-Pi和JS-2-08-Pi;SPF鸡胚购自山东家禽研究所SPF鸡场。
1.2 主要试剂 禽成髓细胞病毒反转录酶(AMV)购自 TOYOBO 公司;200 bp DNA Marker、DNA 凝胶回收试剂盒、dNTP购自TaKaRa公司;Taq DNA聚合酶、T4 DNA连接酶、限制性内切酶EcoR I为MBI公司产品;pCR2.1 载体、Trizol、DH5α 大肠杆菌为 Invitrogen 公司产品;Ampicilin、X-gal、IPTG 购自Sigma公司;DEPC购自BBI公司。
蛋白胨和酵母抽提物购自英国OXOID公司;琼脂粉琼脂糖为西班牙产品;醋酸钠(NaAc)、十二烷基磺酸钠(SDS)系Sigma公司产品;三氯甲烷(氯仿)、异丙醇、异戊醇、乙酸钾、冰乙酸和三羟基甲基氨基甲烷等常规试剂购自中国医药(集团)上海化学试剂公司;饱和酚为南京生物工程公司产品。
1.3 主要仪器 PTC-200型PCR仪、3000XI型电泳仪购自美国BIO-RAD公司;恒温金属水浴锅CHB-100型购自杭州大和热磁电子有限公司;RC232C型紫外分光光度计、台式高速离心机购自德国eppendorf公司;纯水仪Milli-Q低热原型购自美国Millipore公司。
1.4 引物 参照已发表的NDV毒株基因的核苷酸序列,应用Primer Premier5.0设计了两对引物,P1和P2扩增M基因1161 nt至F基因889 nt间长约970 pb的片段,P3和P4扩增F基因862 nt至F基因1700 nt间长约839 bp的片段。引物由上海生工生物工程技术服务有限公司合成,序列为:
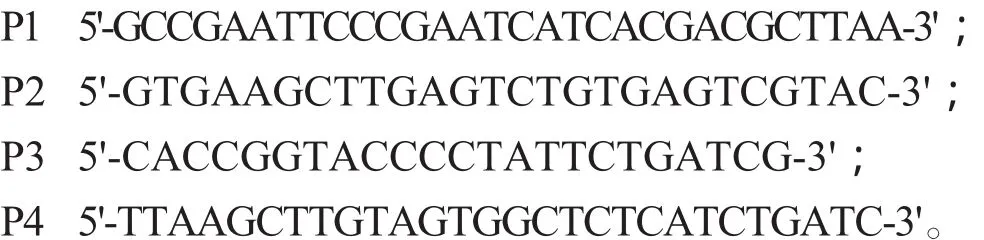
1.5 病毒毒力的测定 按照世界动物卫生组织(OIE)介绍的方法进行[24]。
1.6 病毒RNA的提取和cDNA的合成 NDV毒株的RNA抽提按照Trizol试剂说明书进行。cDNA的合成按照参考文献进行[17]。
1.7 NDV F基因的克隆 F基因的RT-PCR扩增按有关文献报道的方法进行[17]。RT-PCR扩增产物克隆到pGEM-T Easy Vector,转化感受态大肠杆菌,筛选阳性克隆。
1.8 序列测定和推导氨基酸序列分析 筛选出的F基因阳性克隆寄送到上海生工生物工程技术服务有限公司测定核苷酸序列,用DNAStar 5.0版本软件包的软件(DNASTAR Inc.Madison,WI153715,USA)分析F基因的核苷酸序列及推导的氨基酸序列。
1.9 NDV系统进化发生树和限制性内切酶位点分布分析 参考已报道的方法[16-17],根据F基因47nt~420 nt之间的374 bp片段的核苷酸序列绘制NDV系统进化发生树,同时分析F基因334nt~1682nt之间的1349 bp片段上HinfⅠ、BstOⅠ和RsaⅠ三种限制性内切酶位点。
2 结果
2.1 病毒毒力的测定结果 2株新城疫病毒毒株,经毒力检测测得鸡胚平均死亡时间(MDT)分别为48h和46 h,8周龄SPF鸡静脉接种指数(IVPI)分别为2.88和2.91,一日龄雏鸡脑内接种指数(IPCI)分别为1.88和1.89,均判定为强毒株。
2.2 NDV F基因片段的扩增、克隆 该毒株经RT-PCR均扩增出了大小为970 pb和839 bp的特异性条带。
2.3 NDV F基因序列测定和序列分析
2.3.1 NDV F基因序列测定和推导氨基酸序列分析
根据测得的F基因核苷酸序列推导出相应编码氨基酸,蛋白酶裂解位点附近的氨基酸序列为112R(K)RQKR↓F117,符合NDV强毒株的规律[25]。
2.3.2 NDV系统进化发生树和限制性内切酶位点分布
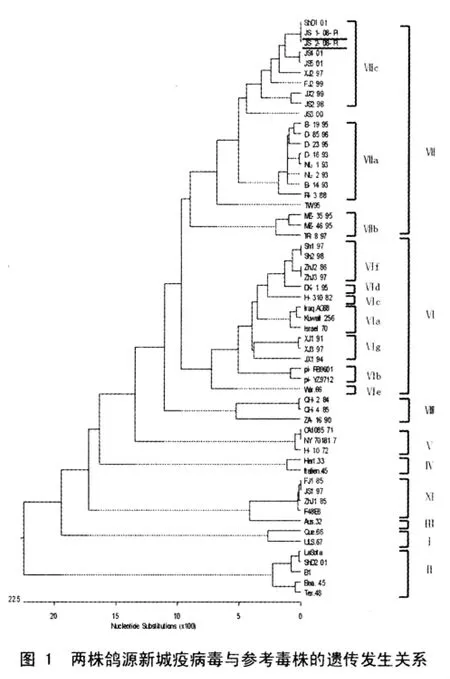
用DNAStar软件对2个鸽源分离株和28个参考毒株的F基因47 nt~420 nt区域进行比较,绘制了系统进化发生树(图1)。结果显示,两个毒株分布在一个分支上,为基因Ⅶ型。
F基因334 nt~1682 nt间三种限制性内切酶(HinfⅠ、BstOⅠ和RsaⅠ)位点分布是基因分型的另一个标准[7-8]。本文中所分离到的鸽源NDV分离株归于Ⅶ型,它们具有特定的限制性内切酶位点分布。在736nt,883nt具有 HinfⅠ位点,而在 1198 nt,1350 nt不具有 HinfⅠ位点,在 752 nt,1116 nt具有 HinfⅠ位点,而在 953 nt,1478 nt、1601 nt不具有 HinfⅠ位点,在 872 nt,973 nt、1249 nt具有 HinfⅠ位点,而在540 nt,683 nt、1593 nt、1625 nt不具有 HinfⅠ位点。
3 讨论
从临床病鸽体内分离到了2株NDV,毒力测定结果表明,2株新城疫病毒毒株的MDT分别为46 h和 48 h,IVPI分别为 2.88和 2.91,IPCI分别为 1.88和1.89,按照OIE推荐的标准为强毒株。F蛋白裂解位点的氨基酸推导序列为112R(K)RQKR↓F117,符合强毒株的分子特征。遗传发生分析表明这两个毒株与同期其它禽群中发生新城疫的毒株亲缘关系很近。
早期研究表明鸽源新城疫病毒抗原性和基因型均有一定的特征,单抗分排谱属于P组[14],基因型属于Ⅵb亚型[16]。1996年我国承办国际信鸽比赛后,排出的病毒使得大范围ND流行。国内不少学者对病原进行了分离和研究。刘文斌等分离到一株鸽源NDV,为基因Ⅵ型[26]。韦平等分离到一株鸽源毒株gxp22也为Ⅵb亚型[27]。刘华雷等分到5株鸽源毒株基因型也为基因Ⅵ型[28]。而本文中分离到的毒株属于Ⅶc亚型。这表明Ⅶc亚型NDV在鸽群中存在,这与以往经典Ⅵb亚型毒株占优势相比出现了新的变化。
对于鸽源新城疫病毒毒力的研究表明,与OIE推荐的标准以及裂解位点氨基酸序列标准不一致。于守平报道了2004年12月份吉林省肉鸽一例NDV,并分离到了一株强毒株[13]。连宏军等从黑龙江发病鸽场分离了两株NDV,经毒力鉴定为强毒株[29]。梁华丽从发病鸽分离出的NDV为弱毒[30]。韦平等分离到的鸽源毒株gxp22裂解位点氨基酸序列为强毒序列,而IVPI为0[27]。刘华雷等报道的5株鸽源毒株裂解位点氨基酸序列为强毒序列,但未进行毒力的测定[28]。李柠等和谢青梅等人的报道也表明鸽源毒株对鸡的致病力差异很大[31-32]。Dortmans J C等人2009年的一篇报道表明鸽源毒株裂解位点氨基酸序列并不总与毒力相关[33]。(2002我们分离的一株鸭源NDV裂解位点氨基酸序列为典型强毒序列,但IVPI为0,未发表。)本文中分离到的两株鸽源新城疫病毒毒力均较高,且裂解位点氨基酸序列为强毒。以上研究都表明我国鸽群中新城疫病毒的毒力和生物学特性存在较大差异。
从发病禽分离到的病毒按照OIE推荐的判定标准鉴定为弱毒的病毒毒株,这说明不同的新城疫毒株对不同宿主的致病力有一定差异。完全用对NDV非常敏感的鸡作为新城疫毒株的唯一试验宿主可能会影响对新城疫病毒毒力正确的判定。
我国鸽的饲养量越来越大,除肉鸽外,信鸽、赛鸽等一些名贵的鸽种也越来越多,随着养殖量增加和规模扩大,疫病防控的难度也随之加大。而由于其它野生禽类,家养禽类可能的带毒排毒状态,会加大养鸽的风险。建立不同禽类新城疫病毒监测体系,对该病的分子流行病学和病原变异进行监控,对于控制新城疫在我国禽群和其它动物群内的传播有着非常重要的意义。因此,对临诊分离到的病毒进行病原体的生物学特征和遗传变异进行监测对该病的防控十分重要。
[1]Alexander D J,Russell P H,Collins M S.Paramyxovirus type 1 infections of racing pigeon:1 characterization of isolated viruses[J].Vet Rec,1984,114(18):444-446.
[2]The Avian Study Group,BSABA.Paramyxovirus infections of pigeons[J].Vet Rec,1984,114(10):232.
[3]Gelb J,Fries P A,Peterson F S.Pathogenicity and cross-protection of pigeon paramyxovirus-1 and Newcastle Disease Virus in young chickens[J].Avi Dis,1987,31(3):601-606.
[4]Lister S A,Alexander D J,Hogg R A.Evidence for the presence of avian paramyxovirus type 1 in feral pigeons in England and Wales[J].Vet Rec,1986,118(17):476-479.
[5]Alexander D J,Parsos G,Marshall R.Infection of fowls with Newcastle Disease virus by food contaminated with pigeon faces[J].Vet Rec,1984,115(23):601-602.
[6]Ballgi-Pordany A,Wehmann E,Herczeg J,et al.Identification and grouping of Newcastle Disease Virus strains by restriction site analysis of a region from the F gene[J].Arch Virol,1996,141(2):243-261.
[7]Alexander D J,Parsons G.Avian paramyxovirus type 1 infections of racing pigeons:2 pathogenicity experiments in pigeons and chickens[J].Vet Rec,1984,114(19):466-469.
[8]Alexander D J.Parsons G.Pathogenicity for chickens of avian paramyxovirus type 1 isolates obtained from pigeons in Great Britain during 1983-85[J].Avi Pathol,1986,15(3):487-493.
[9]李德厚,刘文灿,陈琨,等.从进口种鸽多次检出鸽新城疫病[J]. 中国兽医杂志,1988,14(6):54-55.
[10]佘柏荣.鸽I型副粘病毒概况[J].佛山兽医专科学校学报,1992(2):60-64.
[11]许家强,李淡薇,孔小明.鸽新城疫流行的调查研究[J]. 山西农业大学报,1994,14(2):155-157.
[12]陈昌海,程雷,严建刚.鸽新城疫病毒的分离及其生物学特性测定[J]. 中国预防兽医学报,1999,21(2):98-99.
[13]于守平,胡桂学,董丽娜.鸽新城疫病毒的分离鉴定[J].黑龙江畜牧兽医,2006(8):79-80.
[14]Alexander D J,Manvell R J,Kemp P A,et al.Use of monoclonalantibodiesin the characterization ofavain paramyxovirus type Ⅰ (Newcastle Disease Virus)isolates submitted to an international reference laboratory[J].Avi Pathol,1987,16(4):533-565.
[15]Alexander D J,Russell P H,Parson G,et al.Antigenic and biological characterization of avian paramyxovirus type 1 isolates from pigeonan international collaboration study[J].Avi pathol,1985,14(3):365-376.
[16]Lomniczi B,Wehmann E,Herczeg J,et al.Newcastle Disease out-breaksin recentyearsin western Europe were caused by an old (VI) and a novel genotype(VII)[J].Arch Virol,1998,143(1):49-64.
[17]X F Liu,H Q Wan,X X Ni,et al.Pathotypical and genotypical characterization of strains of Newcastle disease virus isolated from outbreaks in chicken and goose flocks in some regions of China during 1985-2001[J].Arch Virol,2003,148(7):1387-1403.
[18]H L Liu,Z L Wang,C P Song,et al.Characterization of pigeon-origin Newcastle disease virus isolated in China[J].Avi Dis,2006,50(4):636-640.
[19]Meulemans G,van den Berg T P,Decaesstecker M.Evolution of pigeon Newcastle disease virus strains[J].Avi Pathol,2002,31(5):515-519.
[20]Alexander D J,Parsos G,Marshall R.Infection of fowls with Newcastle Disease virus by food contaminated with pigeon faces[J].Vet Rec,1984,115(23):601-602.
[21]陈予召.鸽感染鸽源I型副粘病毒和鸡新城疫病毒的临诊和病理变化的比较 [J].中国兽医杂志,1999,25(5):7-9.
[22]Kim L M,Kin D J,Guzman H,et al.Biological and phylogenetic characterization of pigeon paramyxovirus serotype 1 circulating in wild North American pigeons and doves[J].J Clin Microbiol,2008,46(10):3303-3310.
[23]Abolnik C,Gerdes G. H,Kitching J,et al.Characterization ofpigeon paramyxoviruses(Newcastle disease virus)isolated in South Africa from 2001 to 2006[J].Onderstepoort J Vet Res,2008,75(2):147-152.
[24]世界动物卫生组织.诊断试验和疫苗标准手册[M].农业部畜牧兽医局,译.北京:中国农业科学出版社,1996.
[25]Ballagi-Pordany A,Wehmann E,Herczeg J,etal.Identification and grouping ofNewcastle disease virus strains by restriction site analysis of a region from the F gene[J].Arch Virol,1996,141(2):243-261.
[26]刘文斌,崔尚金,刘立奎,等.东北地区不同宿主NDV分离株的系统发育进化 [J].中国兽医杂志,2005,41(7):3-6.
[27]韦平,韦天超,杨宗维,等.禽Ⅰ型副黏病毒各种禽源分离株毒力及其相关基因的研究[J].病毒学报,2005,21(1):35-42.
[28]刘华雷,周斌,郁斌,等.五株鸽副黏病毒国内分离株F基因片段的克隆和分子特性 [J].病毒学报,2004,20(4):378-381.
[29]连宏军,闫丽辉,刘培欣,等.13株新城疫病毒强毒株的分离鉴定及部分生物学特性 [J].动物医学进展,2006,27(6):64-69.
[30]梁华丽,华炯钢,徐辉,等.不同禽源的禽I型副黏病毒毒力免疫原性及抗原相关性研究[J].畜牧与兽医,2008,40(11):22-26.
[31]李柠,黄引贤,李乃津,等.鸽禽Ⅰ型副黏病毒生物学特性的比较[J].中国畜禽传染病,1994(2):1-5.
[32]谢青梅,毕英佐,曹永长,等.鸽禽Ⅰ型副黏病毒的分离和鉴定[J].中国兽医科技,2000,30(7):8-10.
[33]Dortmans J C,Koch G,Rottier P J,et al.Virulence of pigeon paramyxovirus type 1(PPMV-1)not always correlates with the cleavability of its fusion protein[J].J Gen Virol,2009,90(11):2746-2750.
猜你喜欢
科学大观园(2022年2期)2022-01-23
文萃报·周二版(2021年47期)2021-12-14
农药科学与管理(2019年9期)2019-11-23
农药科学与管理(2019年6期)2019-11-23
中国抗生素杂志(2019年6期)2019-07-06
家禽科学(2019年2期)2019-06-11
食品与生活(2018年7期)2018-09-19
中国畜禽种业(2018年10期)2018-01-19
中国猪业(2017年11期)2017-12-11
中国畜牧兽医文摘(2015年9期)2015-12-29
