
棉铃虫和烟夜蛾生殖行为比较研究
2011-05-10 02:07:26程晓东贾月丽蔡永萍王郑军罗梅浩
河南农业大学学报 2011年3期
程晓东,贾月丽,蔡永萍,王郑军,罗梅浩
(河南农业大学植物保护学院,河南郑州450002)
棉铃虫(Helicoverpa armigera(Hübner))和烟夜蛾(Helicoverpa assulta(Guenée))都是重要的农业害虫,二者同属鳞翅目夜蛾科铃夜蛾属,成、幼虫的形态特征很相似,食性特点、为害症状也较一致,且具有杂交可育性[1],已有研究证明他们属亲缘相近的2个种[2].二者的寄主范围有很大差异,棉铃虫是典型的多食性昆虫,其食料植物在中国有20多科200多种,最喜食锦葵科、茄科和豆科的棉花、玉米、小麦、大豆、烟草、番茄等植物[3]。烟夜蛾是一种寡食性昆虫,寄主植物只有辣椒、烟草等茄科植物[4].一直以来人们都在致力于这2种近缘昆虫之间的差异研究.罗梅浩等[5]通过田间调查和室内饲养研究表明,棉铃虫幼虫体色变化复杂于烟夜蛾;此后又通过在烟草上的生态位研究表明,棉铃虫在烟草上的时间生态位宽度、生态位重叠和竞争系数均大于烟夜蛾[6].张礼生[7]采用 HSG 显带和常规C显带法研究了二者的核型特征,结果显示,烟夜蛾染色体多态性更为显著,而棉铃虫相对稳定.王鸿雷等[8]利用气相色谱法分析了外源不饱和脂肪醇及乙酸酯在棉铃虫和烟夜蛾性信息素腺体中的转化,结果表明棉铃虫性信息素腺体组织具有较高的乙酸酯酶活性,而烟夜蛾性信息素腺体乙酸酯酶活性很低.付晓伟等[9]通过2近缘种夜蛾对低浓度烟草挥发物的触角电位反应研究表明,与棉铃虫相比,烟夜蛾嗅觉神经系统在其寄主定向行为中所起的作用更重要、也更专化.研究生物进化的方法常常是通过鉴定近缘类群,比较性状差异,分析造成差异的遗传基础,进而解释该性状是如何从一个分类群向另一个分类群变化的[10].交配是两性生殖昆虫繁衍后代的重要环节[11],是成虫行为发生改变的转折点,也是具有重要进化意义的过程.鳞翅目昆虫的交配行为具有明显的种特异性,通常呈现出一定的昼夜节律性,并且昼夜节律的微小变化,会对昆虫个体间基因交流产生决定性屏障,从而成为推动昆虫新种形成和种下分化的重要原动力.虽然迄今有关棉铃虫和烟夜蛾生殖生物学特性方面的研究已有很多报道,对其交配、产卵行为也做过简单的描述,但仍缺乏对2种夜蛾生殖行为较为系统的比较研究.充分了解2个近缘种昆虫的交配、产卵等生殖行为,不仅为开展其行为生态学研究提供基础知识,而且对更好预测其种群扩散和物种适应与演化也有一定的参考价值.为此,作者研究了不同蛾龄下棉铃虫和烟夜蛾成虫交配和产卵行为的差异,以期为2个近缘种害虫的综合治理和预测预报提供依据和参考.
1 材料与方法
1.1 供试昆虫及饲养方法
棉铃虫和烟夜蛾均从河南农业大学科教园区采集,在控光(14L:10D)、控温(27±1)℃、控湿(65%~85%)的人工气候箱内饲养,饲养方法同参考文献[12].待化蛹后,把蛹按雌雄分开,成虫羽化后喂以质量浓度为100 g·L-1的蔗糖水.
1.2 棉铃虫和烟夜蛾交配次数的观察
将当日羽化的棉铃虫雌蛾置于600 mL的透明塑料杯中,杯底放一吸水棉球,暗期前1 h每杯中引入1头3日龄的处女雄蛾.当晚如发现交配,次日早上用另1头3日龄的处女雄蛾替换前晚杯中的雄蛾,直至雌蛾死亡.待雌蛾死亡后在显微镜下解剖其交配囊,检查精包有无及数量,以精包的个数代表雌蛾的交配次数[13].本试验设30次重复.雄蛾交配的观察方法同雌蛾,只是其交配次数等于解剖与其交配所有雌蛾精包的累加数.烟夜蛾雌雄蛾交配次数的测定方法同棉铃虫.
1.3 棉铃虫和烟夜蛾交配行为的观察
设当晚羽化的成虫蛾龄为0,第2天的成虫蛾龄为1,依次类推.将当日羽化的棉铃虫或烟夜蛾成虫分别配对置于600 mL的透明塑料杯中,杯底放一吸水棉球,每天定时补充成虫饲料,然后观察其交配行为,观察从羽化当天一直延续到羽化后的第7天.同时,在透明塑料杯的顶部用纱布遮盖,以便成功交配的雌蛾在上产卵.2种群分别观察300对成虫,设3次重复,每重复100对.由于在光照期间,成虫很少有交配行为发生,故观察主要在夜间熄灯后(22:00—08:00)进行.用红光手电筒进行观察,并借助于计时器的帮助,每隔15 min观察1次.在观察过程中,分别记录每对成虫交配开始和结束的时间.
1.4 棉铃虫和烟夜蛾产卵行为的观察
在1.3试验中,如发现雌雄蛾成功交配,每天定期查看纱布上是否有虫卵.一旦发现卵粒马上记下时间,每天用新的纱布替换含卵纱布,把含卵纱布放在明处统计卵粒数,直至雌蛾死亡.把含卵纱布放入培养皿中保湿培养,直至幼虫孵化.然后计算雌蛾寿命、产卵前期、产卵期、产卵量和卵的孵化率.
1.5 数据的统计分析
数据经SPSS17.0统计软件进行单因素方差分析和多重比较,所获数据主要用平均数(mean)±标准误(SE)来表示.百分率数据在比较前先进行反正弦变换.2种夜蛾的种间差异采用t-测验进行比较.
2 结果与分析
2.1 棉铃虫和烟夜蛾的交配能力
在30头棉铃虫雌蛾中,有27头能正常交配,每头雌蛾平均交配1.2次;在30头烟夜蛾雌蛾中,有26头能正常交配,每头雌蛾平均交配1.1次.在30头棉铃虫雄蛾中,有28头能正常交配,交配次数最多的有6次,每头雄蛾平均交配3.0次;在30头烟夜蛾雄蛾中,有26头能正常交配,交配次数最多的有5次,每头雄蛾平均交配2.5次.可见,2种夜蛾之间雌雄蛾的交配能力差异均不显著,但雄蛾的交配次数明显多于雌蛾(表1).

表1 棉铃虫和烟夜蛾的交配能力Table 1 Mating capacities of H.armigera and H.assulta
2.2 不同蛾龄下两种夜蛾的交配率
棉铃虫和烟夜蛾成虫羽化的当天晚上未发生交配,从羽化后的第1天晚上(1日龄)到第7天晚上都可以进行交配(图1).2种夜蛾交配高峰均出现在羽化后的第3天,交配率分别达30%和24%,然后开始下降,7日龄的交配率分别为2%和1%.6日龄前的每日龄棉铃虫的交配率均显著高于烟夜蛾.7日龄前的交配率棉铃虫为93%,烟夜蛾为69%.
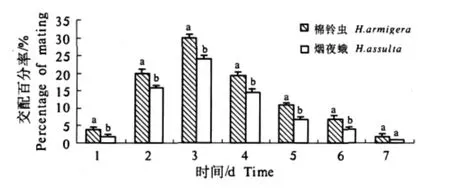
图1 棉铃虫和烟夜蛾羽化后7 d内交配发生的概率分布Fig.1 Distribution of mating rates of H.armigera and H.assulta during the first 7 days after emergence
2.3 2种夜蛾交配的昼夜节律
熄灯后,棉铃虫交配发生较晚,2 h后才有零星交配发生;烟夜蛾交配发生较早,熄灯后1 h就有交配发生.棉铃虫成虫交配高峰在凌晨4:00—6:00,交配发生的概率占全天的54%,到次日早上亮灯时仍有零星发生;而烟夜蛾成虫在光照条件下(白天)不进行交配,交配高峰在凌晨0:00—2:00,交配发生的概率占全天的56%.2种夜蛾的交配高峰出现的时间明显不同(图2).随着黎明的到来,成虫交配的频率又明显降低(图2).
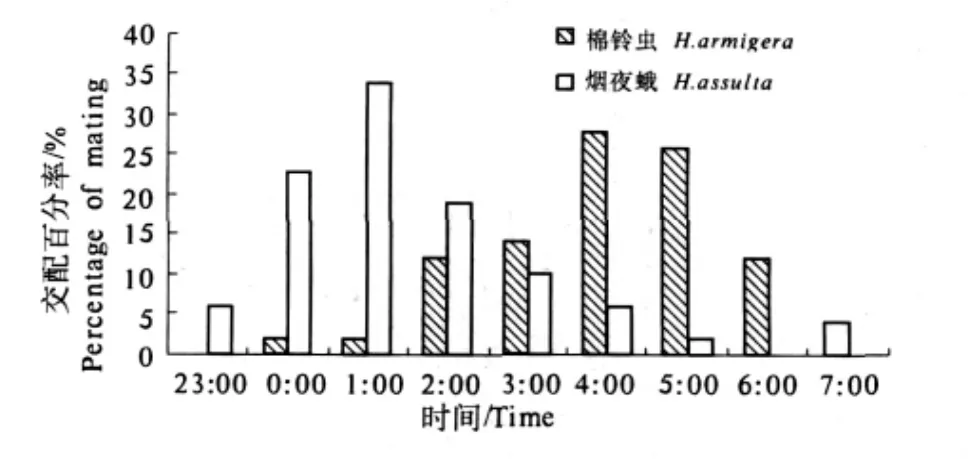
图2 棉铃虫和烟夜蛾交配活动的昼夜节律Fig.2 Circadian rhythm of mating activities of H.armigera and H.assulta
2.4 不同蛾龄棉铃虫和烟夜蛾交配持续时间
棉铃虫一生的交配持续时间平均为84.7 min,烟夜蛾一生的交配持续时间平均为92.8 min.蛾龄对2种夜蛾的交配持续时间具有显著的影响,低日龄成虫的交配持续时间较短,随着蛾龄的增加,平均交配持续时间逐渐延长(图3),1日龄棉铃虫和烟夜蛾平均交配持续时间分别为48.8,62.6 min,而7日龄则平均分别为105,116.7 min,相当于1日龄的2倍.且不同蛾龄下烟夜蛾的交配持续时间均长于棉铃虫,两者之间的差异达显著水平.
2.5 不同蛾龄棉铃虫和烟夜蛾的产卵行为

图3 不同蛾龄棉铃虫和烟夜蛾的交配持续时间Fig.3 Mating duration of H.armigera and H.assulta in different age
由表2,表3结果可以看出,1日龄交配的棉铃虫和烟夜蛾产卵前期分别为2.5,3.6 d;7日龄交配的2种夜蛾产卵前期分别为8.3和9.3 d;棉铃虫和烟夜蛾不同交配蛾龄的产卵前期之间均存在显著差异;同一蛾龄2种夜蛾交配前期之间亦存在显著差异,4日龄前两者的差异达极显著水平.随着交配蛾龄的增加,2种夜蛾的产卵前期也显著延长,与蛾龄之间存在显著的正相关.不同交配蛾龄棉铃虫的寿命均长于烟夜蛾,且4日龄前两者之间差异性显著;同样,随着交配蛾龄的增加,2种夜蛾的寿命也显著延长,与蛾龄之间亦存在显著的正相关.
棉铃虫和烟夜蛾的产卵期在3日龄前差异显著,4日龄后无显著差异,产卵期与蛾龄之间存在显著的负相关.棉铃虫和烟夜蛾的产卵量在3日龄前相对较高,并且5日龄前棉铃虫的产卵量均显著高于烟夜蛾;产卵量亦与蛾龄之间存在显著的负相关.1日龄棉铃虫和烟夜蛾卵的孵化率分别为74.2%和62.3%,7日龄卵的孵化率分别为44.6%和21.8%,可见,随着蛾龄的增加,2种夜蛾所产卵的孵化率降低,且不同蛾龄棉铃虫卵的孵化率均显著高于烟夜蛾;同样,卵孵化率与蛾龄之间存在显著的负相关.

3 结论与讨论
本研究结果表明,2种夜蛾雌雄蛾交配能力之间均无显著差异.关于昆虫多次交配的动力和适应意义已有许多相关研究,鉴于雌雄两性在配子投入上的差异,雄虫多次交配的益处显而易见.雄虫是竞争的性别,它的生殖成功受与其交配的配偶数量的限制,与它交配的配偶数量越多,表明卵受精的机会越大,产生较多后代的可能性越高,从而获得高的适应性[14].
棉铃虫和烟夜蛾的交配率均表现为3日龄时最高,分别为30%和24%,且6日龄前棉铃虫的交配率均显著高于烟夜蛾.与李国清等[15]报道的交配高峰在4日龄略有不同;3日龄后交配率开始下降,7日龄的交配率分别为2%和1%,说明随着蛾龄的增加,成虫性器官先是慢慢发育成熟,然后再退化.在其他蛾类昆虫中,如棉红铃虫(Pectinophora gossypiella)和欧洲葡萄蛾(Lobesia botrana)也有类似报道[16,17].另外,棉铃虫和烟夜蛾一生的交配持续时间平均为84.7和92.8 min,不同蛾龄下2种夜蛾的交配持续时间存在显著差异;这可能是因为棉铃虫的活动性较强,烟夜蛾则较弱,烟夜蛾的雄虫需要延长交配时间才能完成精子的转移过程,或者是雄虫需要通过延长交配时间来转移更多的精子或其他附腺物质.并且随着蛾龄的增加,其交配持续时间逐渐延长.这一结果与甜菜夜蛾(Spodoptera exigu)和大猿叶虫(Colaphellus bowringi Baly)的研究结果一致[18,19].
棉铃虫和烟夜蛾成虫交配高峰发生的时间明显不同.造成这种差异的原因可能是性信息素释放节律不同[21]或寄主植物的影响[22].同一物种的各类群间因交配节律不同而导致生殖隔离的事例很多,如草地贪夜蛾(Spodoptrra fngiperda)的玉米类群与水稻类群[23]、二化螟(Chilo suppressalis)的水稻类群和茭白类群[24],其交尾时间的差异是导致生殖隔离的重要原因之一.棉铃虫与烟夜蛾虽不是同一种昆虫,但是否会因交配节律的差异而对相互间的交配产生影响,值得深入研究.
成功的繁殖是害虫为害的基础.了解交配后雌虫生理和行为的变化以及引起这些变化的刺激因子具有重要的理论和实用价值.产卵行为研究结果表明:随着交配蛾龄的增加,雌蛾产卵量下降,卵孵化率降低,产卵期缩短,与蛾龄之间存在显著的负相关;而产卵前期和寿命逐渐延长,与蛾龄之间存在显著的正相关,这些结果在应用性信息素防治2种害虫的实践中无疑具有特别重要的意义.不同交配蛾龄棉铃虫的产卵前期均短于烟夜蛾,且二者差异显著;产卵前期的差异从一定程度上反映了卵成熟的进程以及卵巢内卵黄原蛋白含量的不同,涉及到的代谢途径和具体功能仍需进一步研究.而不同蛾龄棉铃虫的寿命均略长于烟夜蛾,且5日龄前二者有显著差异.随着交配蛾龄的增加,两种夜蛾的寿命也显著延长,与蛾龄也存在显著的正相关;这些与烟芽夜蛾(Heliothis virescens)和水稻二化螟Chilo suppressal)等鳞翅目昆虫的研究结果相似[25,26].不同蛾龄的棉铃虫卵孵化率均高于烟夜蛾,且二者存在显著差异;但卵孵化率与蛾龄存在显著负相关,雌蛾蛾龄变化引起的卵孵化率的下降可能是由于未成功授精卵的增多.随着蛾龄的增加,未交配雌蛾退化的卵母细胞沉积物也增加,当雌雄蛾最后交配时,这些沉积物能阻止精包运送到交配囊[27].
5日龄前棉铃虫和烟夜蛾的产卵量存在显著差异,且棉铃虫的产卵量远大于烟夜蛾.关于2种近缘种夜蛾产卵量相差如此之大的原因可能是由于卵巢分泌蜕皮甾醇[28]和合成卵黄蛋白表达量的不同[29],或同一发育时期卵巢、精巢等性器官发育程度的不同以及生殖系统内糖类、蛋白质等营养物质含量的不同造成的,这些都有待进一步研究.随着成虫蛾龄的增加,2种夜蛾产卵量减少的原因可能是在交尾过程中雌蛾体内生殖附腺分泌物和荷尔蒙数量和品质的降低或者无这些分泌物,或者是雌蛾体内退化卵母细胞的残留物阻止雄蛾精包运送到交配囊内导致交配后雌蛾体内精包数的下降[30].因此,在田间应用信息素防治2个近缘种害虫时,可以干扰或延迟其交配,这样就可降低雌蛾的产卵量和卵孵化率,实现降低田间种群数量的目的.
[1] WANG C Z.Interpretation of the biological species concept from interspecific hybridization of two Helicoverpa species[J].Chinese Science Bulletin,2007,52(2):284-286.
[2] 王琛柱.从棉铃虫和烟青虫的种间杂交理解生物学物种概念[J].科学通报,2006,56(21):2573-2575.
[3] 郭予元.棉铃虫的研究[M].北京:中国农业出版社,1998:22-24.
[4] 马继盛,罗梅浩,郭线茹,等.中国烟草昆虫[M].北京:科学出版社,2007.
[5] 罗梅浩,郭线茹,张宏亮,等.烟田烟青虫和棉铃虫幼虫体色变化及遗传规律初步研究[J].河南农业大学学报,1999,33(3):263-266.
[6] 罗梅浩,郭线茹,郑晓军,等.烟青虫和棉铃虫在烟草上的生态位及其种间竞争[J].中国烟草学报,2002,8(4):34-37.
[7] 张礼生.棉铃虫与烟青虫细胞遗传性研究[D].北京:中国农业大学,2004.
[8] 王鸿雷,赵成华,闫云花,等.棉铃虫和烟青虫性信息素腺体脂肪醇和乙酸酯转化的比较研究[J].昆虫学报,2008,51(9):895-901.
[9] 付晓伟,郭线茹,罗梅浩,等.两近缘种夜蛾对低浓度烟草挥发物的触角电位反应[J].生态学报,2009,29(6):2962-2970.
[10] FOSTER S P,MUGGLESTON S J,LÖFSTEDT C.A genetic study on pheromone communication in two Ctenopseustis moths[C]//CARDÉ R T,MINKS A K.Insect Pheromone Research:New Directions.New York:Chapman & Hall,1997:514-524.
[11] MIYATAKE T.Correlated responses to selection for developmental period in Bactrocera cucurbitae(Diptera:Tephritidae):Time of mating and daily activity rhythms[J].Behavior Genetics,1997,27(5):489-498.
[12]罗梅浩,薛伟伟,刘晓光,等.不同烟草品种对烟实夜蛾和棉铃虫产卵引诱作用的研究[J].河南农业大学学报,2006,40(2):198-200,204.
[13]王厚振,华尧楠,牟吉元.棉铃虫预测预报与综合治理[M].北京:中国农业出版社,1999:23-29.
[14]刘兴平,彭接辉,何海敏,等.多次交配对昆虫适应性的影响[J].江西农业大学学报,2008,30(4):592-600.
[15]李国清,陈长琨,韩召军,等.棉铃虫蛾性成熟与交配习性的研究[J].南京农业大学学报,1998,21(2):42-46
[16] LINGREN P D,WARNER W B,HENNEBERRY T J.Influence of delayed mating on egg production,egg viability,mating and longevity of female pink bollworm(Lepidoptera:Gelechiidae)[J].Environmental Entomology,1988,17(1):86-89.
[17] TORRES-VILA L M,RODRÍGUEZ-MOLINA M C,STOCKEL J.Delayed mating reduces reproductive output of female European grapevine moth,Lobesia botrana[J].Bulletin of Entomology Research,2002,92(3):241-249.
[18]罗礼智,曹卫菊,钱 坤,等.甜菜夜蛾交配行为和能力[J].昆虫学报,2003,46(4):494-499.
[19]蒋金炜,丁识伯,张艳民,等.人工饲料对棉铃虫生长发育和繁殖力的影响[J].河南农业大学学报,2010,44(1):78-85.
[20]刘兴平,何海敏,匡先钜,等.影响大猿叶虫交配持续时间的因素[J].昆虫学报,2010,53(5):549-554.
[21] KANNO H.Seasonal variation in periodicity of mating behaviour in the rice stem borer,Chilo suppressalis(Walker)(Lepidoptera:Pyralidae)[J].Bulletin of Entomological Research,1981,71(4):631-637.
[22] DICKENS J C,SMITH J W,LIGHT D M.Green leaf volatiles enhance sex attractant pheromone of the tobacco budworm,Heliothis virescens(Lep.:Noctuidae)[J].Chemoecology,1993,4(3):175-177.
[23] PASHLEY D P,HAMMOND A M,HARDY T N.Reproductive isolating mechanisms in fall armyworm host strains[J].Annals of the Entomological Society of A-merica,1992,85(4):400-405.
[24]孙丽娟,戴华国,衣维贤,等.二化螟水稻类群与茭白类群成虫羽化节律和交配节律研究[J].昆虫知识,2002,39(6):421-423.
[25] HENNEBERRY T J,CLAYTON T E.Tobacco budworm moths(Lepidoptera:Noctuidae):Effect of time of emergence,male age,and frequency of mating on sperm transfer and egg viability[J].Journal of Economic Entomology,1985,78(2):379-382.
[26]焦晓国,宣维健,盛承发.水稻二化螟的交配行为[J].生态学报,2006,26(4):1110-1116.
[27] LUM P T M.Oocyte degeneration in Plodia interpunctella H.ubner and Cadra cautella(Walker)(Lepidoptera:Pyralidae):influence of temperature and humidity[J].Environmental Entomology,1983,12:1539-1541.
[28]王荫长,李国清,陈长琨,等.昆虫生物化学[M].北京:中国农业出版社,2001:405-409.
[29]戈林泉,吴进才.昆虫卵黄蛋白及其激素调控的研究进展[J].昆虫知识,2010,47(2):236-246.
[30] FANGNENG H,BHADRIRAJU S.Effects of delayed mating on reproductive performance of Plodia interpunctella(Hübner)(Lepidoptera:Pyralidae)[J].Journal of Stored Products Research,2003,39:53-63.
猜你喜欢
四川蚕业(2021年1期)2021-02-12 02:03:12
今日农业(2020年17期)2020-12-15 12:34:28
今日农业(2020年13期)2020-12-15 09:08:51
昆虫学报(2020年6期)2020-08-06 06:42:52
今日农业(2019年13期)2019-08-12 07:59:08
新疆农垦科技(2016年2期)2016-08-21 13:50:22
现代农业(2016年5期)2016-02-28 18:42:35
现代农业(2016年4期)2016-02-28 18:42:09
中学生物学(2016年8期)2016-01-18 09:08:26
中国马铃薯(2015年3期)2015-12-19 08:03:56
