
干旱和灌溉条件下少花蒺藜草分株生物量分配特征
2011-04-25 09:43张志新
草业科学 2011年2期
张志新,田 迅
(1.兰州大学草地农业科技学院 甘肃草原生态研究所,甘肃 兰州730020;2.内蒙古民族大学生命科学学院,内蒙古 通辽 028043)
少花蒺藜草(Cenchruspauciflorus)俗名刺蒺藜草、草狗子、草蒺藜、洋蒺藜狗子。禾本科蒺藜草属,一年生草本植物[1-2]。总状花序顶生,穗包藏在由多数不育小枝形成的球形刺苞内,每个刺苞含2 粒种子,少数含3粒种子。茎秆扁圆形,基部屈膝或横卧于地面而于节上生根,且分蘖新枝条[3]。原产于北美洲及热带沿海地区,耐旱、抗寒、耐贫瘠、耐修剪、抗病虫害,具有极强的适应性和竞争力,适于在沙质土壤中生长[4-6]。常生长于低海拔地区的耕地、荒地、牧场、路旁、草原、沙丘、河岸和海滨沙地等。繁殖方式以有性繁殖为主[7-8]。少花蒺藜草繁殖能力强、传播途径多。作为生态入侵种,几年之内便可形成优势种[5],与其他植物争夺水分、土壤养分。由于种子的倒刺会对牲畜造成伤害,该草种一般对农牧业生产具有危害作用。但其适口性良好且适于在沙地生长,故有一定的饲用价值,并可用于防风固沙,对沙漠化治理和生态恢复建设具有重要的意义。
植物的表型可塑性是植物适应异质环境的生态表征。张慧荣和杨持[9]研究了羊草(Leymuschinensis)在不同条件下的可塑性变化,发现干旱条件下羊草的根茎和茎节数都有所增加。王静等[10]、张学勇等[11]通过研究放牧干扰下冷蒿(Artemisiafrigida)和2个生境结缕草(Zoysiajaponica)无性系种群构件特征得出,生物构件的数量特征反映了植物的适应机制,表型可塑性是植物与异质环境长期相互作用的结果。目前,有关少花蒺藜草的研究主要集中在对农牧业危害的调查及防控方面[2,5,12],还没有很好的方法能彻底防治。有关少花蒺藜草生物构件的数量特征及其对环境的适应机理报道较少。本研究以科尔沁沙地不同水分条件下的少花蒺藜草为研究对象,从构件水平初步探讨其生长分配的可塑性变化和规律,为揭示该植物适应环境变化的对策和合理利用及控制提供科学依据。
1 材料与方法
1.1研究区自然概况 研究区位于通辽市南郊南沙坨子。地理位置43°36′ N, 122°22′ E,海拔178.5 m;属典型温带大陆性季风气候地带。春季干旱多风,夏季炎热,秋季凉爽,冬季干冷;降水多集中在6-8月。年降水量350~450 mm,无霜期150 d,年平均气温5.8 ℃。研究样地地势起伏,少花蒺藜草呈块状和带状分布,在个别生境中为单一优势种。多在车辙中分布,也偶见于道路两旁,生长状况各异。研究样地植物多以旱生为主,多年生植物主要有小叶锦鸡儿(Caraganamicrophylla)、麻黄(Ephedraequisetina)、差巴嘎蒿(A.halodendron)、芦苇(Phragmitescommunis)等[13-14],与少花蒺藜草相伴而生。个别地段有人工林分布。植物盖度为30%~50%。土壤类型为沙质土壤,部分地段有灌溉。
1.2取样方法 2007年10月15日,将研究区区分为干旱和灌溉2个微生境,对成熟期少花蒺藜草进行取样。干旱微生境无灌溉。灌溉微生境地势较低,并自7月15日起每10 d灌水1次,直至取样。对于每一生境,设置样方随机对少花蒺藜草种群进行取样。样方大小为1 m×1 m,共设4次重复。每重复随机选取9个植株。将所选植株连同根部一并挖出,轻轻抖落根部所附沙土,保持植株的完整,分别放入干净塑料袋内,标记、封口,带回实验室进行测定。
1.3测定与方法 将少花蒺藜草植株样品的根部剪下,清洗干净,吸干水分。测定每一样品地上部分的长度,将样品依叶片、叶鞘、茎和果穗分开,用分析天平(精确度为0.000 1 g)称量,分别装入纸袋中并做好标记。然后将所有构件样品置于80 ℃烘箱中烘48 h,称烘干质量。分别计算根、叶片,叶鞘、茎和果穗构件的生物量占植株总生物量的比例。
1.4数据处理与分析 应用Excel 2003软件进行数据统计与绘图。采用SPSS 15.0(SPSS Inc.,USA)软件对2个微生境下各构件生物量之间的差异进行比较和独立样本T检验;构件生物量与分株生物量之间的关系,株高与各构件、分株生物量之间的关系以及各构件之间的关系均采用相关和回归分析。
2 结果与分析
2.1分株高度、根和穗生物量特征 结果显示,2个生境条件下少花蒺藜草的分株高度和穗生物量均存在显著差异(P<0.05) (表1)。灌溉条件下,植株高度为41.20 cm,明显高于干旱条件的33.70 cm;果穗生物量是干旱条件下的1.8倍。表明少花蒺藜草对环境差异具有显著的适应反应。
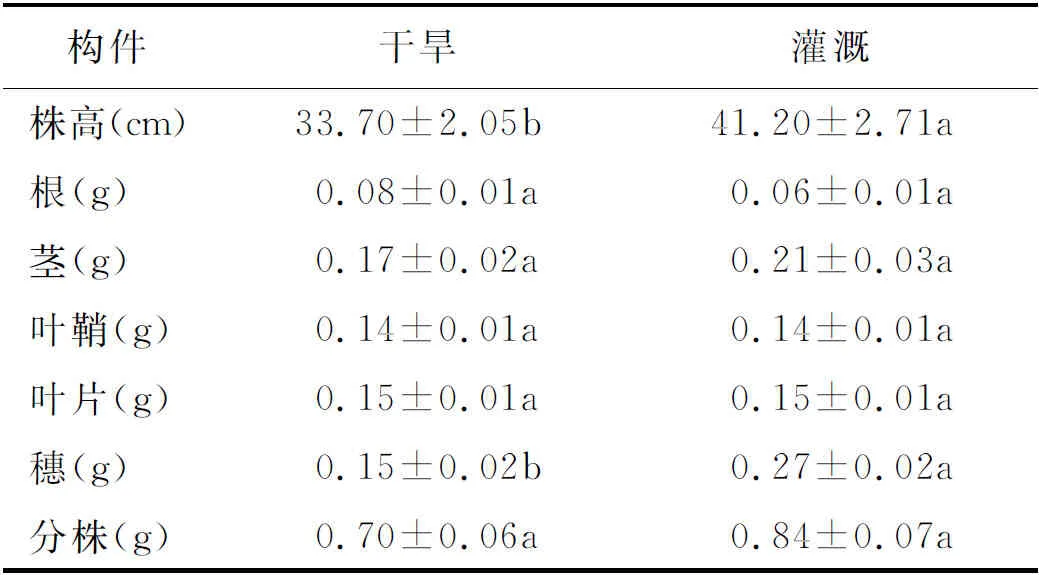
表1 不同条件下少花蒺藜草种群分株的数量特征
2.2构件生物量与分株生物量的关系 相关分析结果表明,2种条件下,根、茎、叶片、叶鞘生物量均与分株生物量呈极显著的正相关(P<0.01),灌溉条件中相关系数(r)依次为:0.777、0.931、0.928和0.921;干旱条件中相关系数(r)依次为0.561、0.908、0.835和0.947。
回归分析及曲线拟合的结果表明,根、茎、叶片、叶鞘生物量与分株生物量的关系符合y=axb幂函数的变化规律。在灌溉条件下,各方程的幂值(b)均高于干旱条件下相应幂值,叶片生物量的b值差异最大。表明随着分株生物量的增加,灌溉条件下各构件的生物量增长速度均高于干旱条件(图1)。
2.3构件生物量与植株高度的关系 相关分析的结果表明,少花蒺藜草在不同生境条件下,根、茎、叶片、叶鞘生物量和分株生物量5个指标与分株高度均呈极显著的正相关(P<0.01)关系,且符合y=axb幂函数的变化规律。在灌溉条件下,方程的幂值(b)均高于干旱条件中相应幂值。表明随着分株高度的增加,灌溉条件下各构件的生物量增长速度均高于干旱条件(表2)。
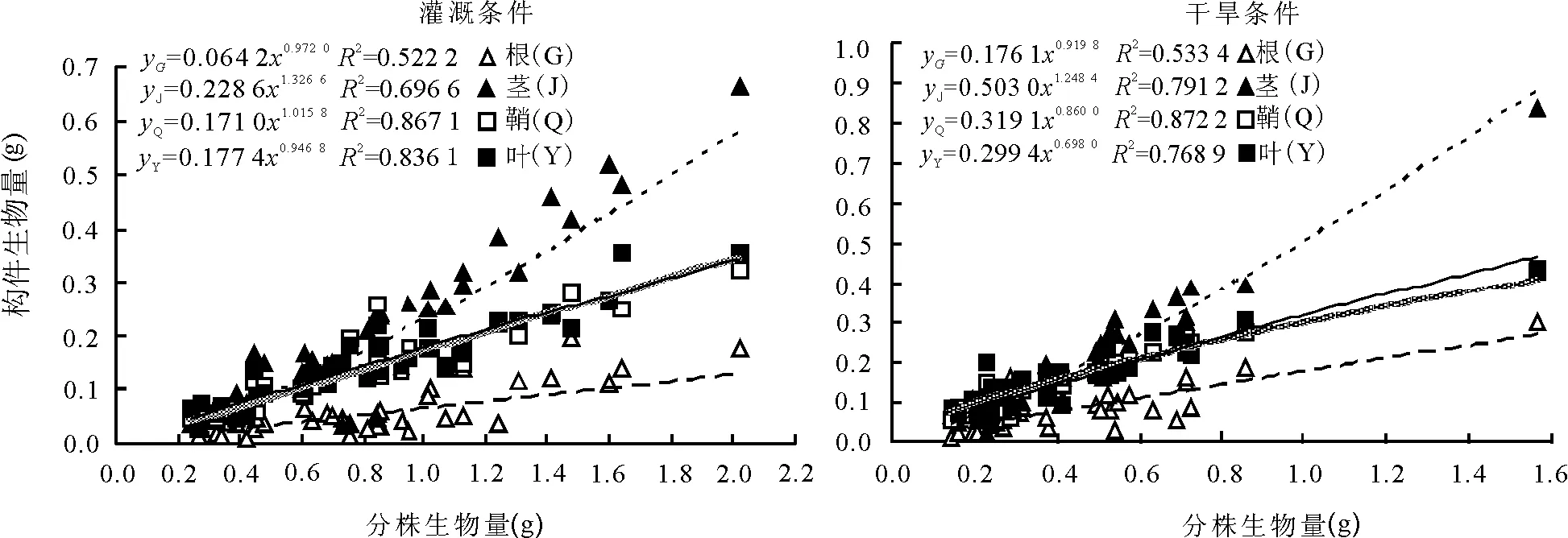
图1 2种条件下少花蒺藜草分株生物量与各构件生物量的关系
2.4构件生物量之间的关系 相关分析结果显示,灌溉条件下,根、茎、叶片和叶鞘生物量之间均呈极显著的正相关(P<0.01);干旱条件下,根与叶鞘生物量、茎生物量、叶片生物量;茎与叶片生物量、叶鞘生物量;叶片与叶鞘生物量之间呈显著的正相关(P<0.01)。以上结果表明,构件生长是相互关联的。
回归分析以及曲线拟合的结果表明,各构件之间的关系符合线性函数模型y=ax+b。拟合的线性方程参数见表3。
表3数据显示,干旱条件下叶鞘、茎、叶片生物量基于根生物量的增长速率小于灌溉条件;而叶鞘、叶片生物量基于茎生物量的变化大于灌溉条件,叶片生物量基于叶鞘生物量的增长速率小于灌溉条件。
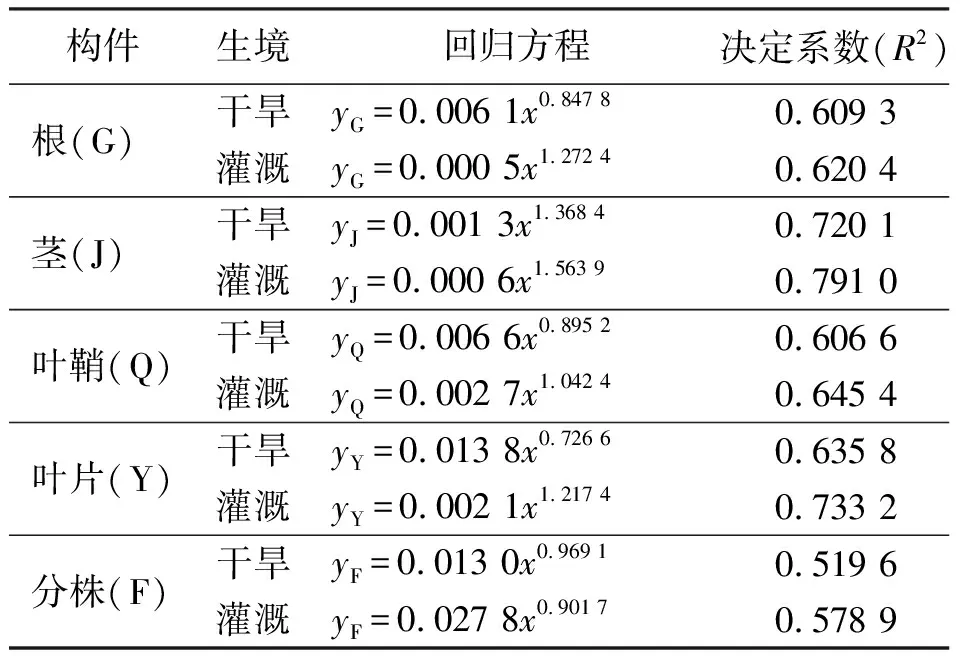
表2 不同条件下少花蒺藜草种群分株高度(y)与构件(x)生物量的关系
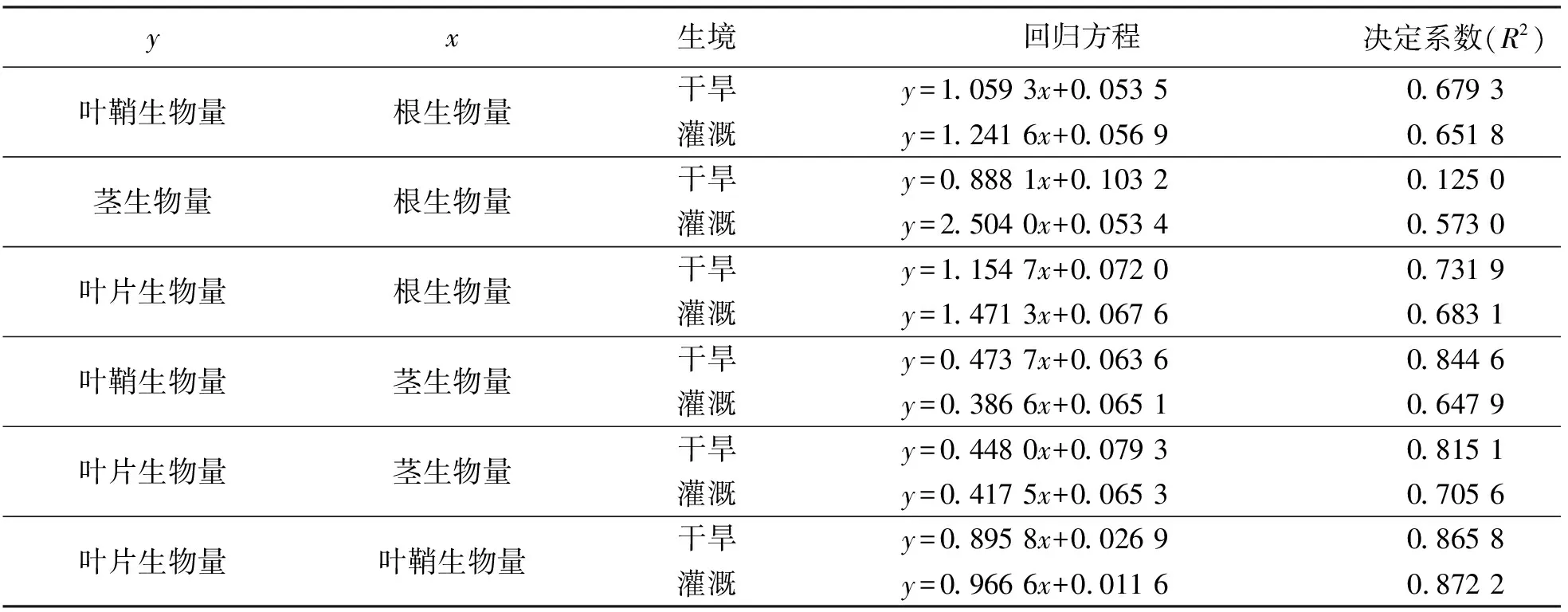
表3 不同条件下少花蒺藜草种群构件生物量之间的关系
3 讨论
结果显示,少花蒺藜草的生长和繁殖性能在灌溉和干旱条件下表现出一定的差异。干旱条件下,少花蒺藜草分株高度降低、穗生物量变小,在物质分配上表现出的是存活对策。灌溉条件下,少花蒺藜草分株高度增加、穗生物量变大,在物质分配上表现出的是竞争和繁殖生长[15]。这与王齐等[16]对水分胁迫下结缕草种群进行研究后得出的株高与生物量随水分胁迫的增加而降低的结论一致。少花蒺藜草在2个生境条件下所表现的差异,体现出少花蒺藜草应对不同环境条件的生态适应对策特征。
少花蒺藜草根、茎、叶片、叶鞘生物量与分株生物量紧密关联。在灌溉条件下,随着分株生物量的增加,各构件的生物量增长速度均高于干旱条件,且以叶片的增长速度为最大;随着分株高度的增加,各构件的生物量增长速度均高于干旱条件。这一结果表明少花蒺藜草在灌溉条件下的物质分配原则是优先建造叶器官与其他植物进行竞争。2种生境条件下少花蒺藜草各构件生物量之间也是紧密关联的。有所不同的是,基于根的生长,在干旱条件下少花蒺藜草向叶鞘、茎、叶片生长的物质分配要小于灌溉条件;基于叶鞘的生长向叶片生长的物质分配要小于灌溉条件,基于茎的生长向叶片和叶鞘生长的物质分配要大于灌溉条件。这些物质分配特征从不同侧面反映了少花蒺藜草适应干旱环境的生态响应机制。
通过本研究的结果和分析,可以看出在物质分配上,干旱条件下少花蒺藜草表现出的是存活对策;灌溉条件下表现出的是竞争和繁殖生长。
[1] Umberto Q.CRC World Dictionary of Grasses[M].Boca Raton,FL:Taylor & Francis,2006:442-443.
[2] 可欣,张秀玲,刘柏,等.彰武县少花蒺藜草发生情况及防除技术[J].杂粮作物,2006,26(1):39-40.
[3] 唐昆.外来入侵生物少花蒺藜草[J].湖南农业,2006(5):16.
[4] Kucera C L.The Grasses of Missouri[M].Missouri:University of Missouri Press,1961:61-62.
[5] 杜广明,曹凤芹,刘文斌,等. 辽宁省的少花蒺藜草及其危害[J].中国草地,1995(3):71-73.
[6] 邱月,庄武, 曲波,等.少花蒺藜草辽宁省分布现状、存在问题及防控建议[J].农业环境与发展, 2009(3):56-57.
[7] 王巍,韩志松.外来入侵生物——少花蒺藜草在辽宁地区的危害与分布[J].草业科学,2005,22(7):63-64.
[8] McCarty L B,Everest J W,Hall D W,etal.Color Atlas of Turfgrass Weeds[M].Chelsea:AnnArbor Press,2001:269-270.
[9] 张慧荣,杨持.不同生境条件下羊草种群构件的可塑性变化[J].内蒙古大学学报(自然科学版),2008,39(5):321-324.
[10] 王静,杨持,王铁娟,等.冷蒿(Artemisiafrigida)种群在放牧干扰下构件的变化[J].生态学报,2006,26(3):960-965.
[11] 张学勇,杨允菲,邵奎龙,等.辽东半岛不同生境结缕草无性系种群构件生物量结构[J].草业科学,2006,23(4):78-81.
[12] 赵艳,吕林有,王巍,等.苜蓿不同播种行距对防控少花蒺藜草的效果[J].草业科学,2010,27(4):78-81.
[13] 李明,蒋德明,押田敏雄,等.科尔沁沙地人工固沙群落草本植物种群生态位特征[J].草业科学,2009,26(8):10-16.
[14] 左小安,赵哈林,赵学勇,等.科尔沁沙地不同恢复年限退化植被的物种多样性[J].草业学报, 2009,18(4):9-16.
[15] 周婵,杨允菲.松嫩平原两个生态型羊草叶构件异速生长规律[J].草业学报,2006,15(5):77-78.
[16] 王齐,孙吉雄,安渊,等.水分胁迫对结缕草种群特征和生理特性的影响[J].草业学报, 2009,18(2):33-38.
猜你喜欢
亚太传统医药(2021年9期)2021-10-26
落叶果树(2021年6期)2021-02-12
文艺生活·中旬刊(2020年8期)2020-11-23
浙江中医杂志(2019年5期)2019-01-06
农机化研究(2018年2期)2018-06-05
读者·校园版(2018年11期)2018-05-15
植物保护(2016年1期)2016-09-14
体育科技(2016年2期)2016-02-28
西华师范大学学报(自然科学版)(2015年3期)2015-02-27
应用海洋学学报(2014年2期)2014-11-26
