
川西北不同退化程度高山草甸和亚高山草甸的植被特征
2011-04-25 09:41:48干友民罗元佳冯国英
草业科学 2011年6期
纪 磊,干友民,罗元佳,成 平,冯国英,马 超
(四川农业大学草业科学系,四川 雅安 625014)
青藏高原是我国主要的畜牧业基地之一,草地资源丰富。然而,目前青藏高原草地退化严重,据统计,约有0.45×108hm2退化草地,约占青藏高原草地面积的1/3[1]。川西北地区处于青藏高原东南缘,属于海拔较高地区,高山草甸与亚高山草甸是该地区主要的植被类型之一,两者普遍处于放牧退化过程中,而植被退化先于土壤退化,认识放牧干扰下草地退化的植被特征是采取合理的管理措施以防止草地退化、保证草地畜牧业可持续发展的基本前提。本研究分别以四川甘孜州理塘县境内的高山草甸与阿坝州红原县境内的亚高山草甸退化草地为研究对象,对其退化演替过程中的植被特征进行了对比研究,以揭示两者退化过程中植被特征变化的相似性与相异性。
1 材料与方法
1.1研究区概况 高山草甸退化草地研究区选在四川省甘孜藏族自治州理塘县境内。地理位置为29°54′~30°14′ N, 99°52′~100°21′ E,平均海拔4 020 m,属高原寒温带季风气候,多年平均气温3 ℃,最高26.8 ℃,最低-30.5 ℃;年平均降水量725.8 mm;相对湿度70%;年平均日照时数2 500~2 650 h。土壤以高山草甸土和亚高山草甸土为主。高山嵩草(Kobresiapygmaea)和四川嵩草(K.setchwanensis)为植被群落主要建群种,伴生种有羊茅(Festucaovina)、西藏嵩草(K.tibetica)、异针茅(Stipaaliena)、垂穗披碱草(Elymusnutans)等[2]。
亚高山草甸退化草地研究区选在四川省阿坝藏族羌族自治州红原县境内。地理位置为31°51′~33°19′ N,101°51′~103°23′ E,平均海拔3 500 m左右。属大陆性高原温带季风气候,年均温为0.6~1.1 ℃,年均降水量为650~730 mm,≥10 ℃年积温865 ℃·d。土壤主要为亚高山草甸土和草甸潮土,植被类型主要为亚高山草甸,主要优势植物有垂穗披碱草、四川嵩草、西藏嵩草、珠芽蓼(Polygonumviviparnm)等[2]。
1.2研究方法
1.2.1植被调查 样地设置采用时空替换法,即利用空间的变换来代替时间的变换,用空间上不同草地放牧退化程度来研究草地在时间上的退化序列[3-4]。通过实地走访与调查,根据不同放牧强度,结合红原县与理塘县国家天然草地退牧还草工程点位置、历史调查资料、牧民居住点位置以及草地放牧时间等因素,在高山草甸与亚高山草甸内分别按照植被盖度及地表裸露程度选取轻度退化(light degradation,LD,地表少量裸露,植被盖度75%~90%)、中度退化(medium degradation,MD,大量裸斑,植被盖度65%~75%)、重度退化(heavy degradation,HD,裸地或少量毒害草,植被盖度<60%)及无明显退化(undegraded,UD,几乎无裸斑,植被盖度>90%)4个夏季天然放牧草地作为研究样地,其中无明显退化样地从围栏5年以上至今无放牧干扰的草地内选取,各样地面积大于1 hm2。各样地远离公路,以减小交通运输对草地的干扰。
2009年7月采用样方法在上述各个样地内分别随机选择5个样方(1 m×1 m)进行植物群落调查,共计130个样方,调查指标包括样方内所有植物种分种盖度、高度、多度、分种地上现存量和频度,以及群落的高度和盖度。
1.2.2数据处理 根据测定结果计算物种优势度:SDR5=(相对高度+相对多度+相对盖度+相对频度+相对质量)/5。
生物多样性指数的计算包括:
S=n
(1)
物种多样性指数(Shannon-Wiener Index):
(2)
均匀度指数(Pielou Index)[5]:
(3)
式中,n为出现在样方中的物种数目,Pi为第i个种的相对优势度(SDR5i/100)。
以上数据的统计分析采用Excel和SPSS软件,图中数据结果均以平均数±标准差表示。
2 结果与分析
2.1物种丰富度与优势种变化 不同退化程度的高山草甸草地物种丰富度变化从无明显退化(UD)到重度退化(HD)呈高-低-高趋势,与亚高山草甸(低-高-低)呈相反趋势,说明放牧干扰对两类退化草地物种丰富度的影响具有相异性(图1)。高山草甸退化演替上草地丰富度值变化范围为9.56~15.00种/m2(轻度退化最低),亚高山草甸变化范围为11.80~25.08种/m2(轻度退化最高)。草地群落则由高大丛生的嵩草群落向低矮耐牧的匍匐型、莲座状植物组成的杂类草群落演变,其间以直立多年生杂类草群落为过渡。随着放牧强度增加,高山草甸某些物种如四川嵩草从群落中消失,而另一些植物,如高山嵩草则通过形态变化或耐践踏适应放牧等变化而得以生存。
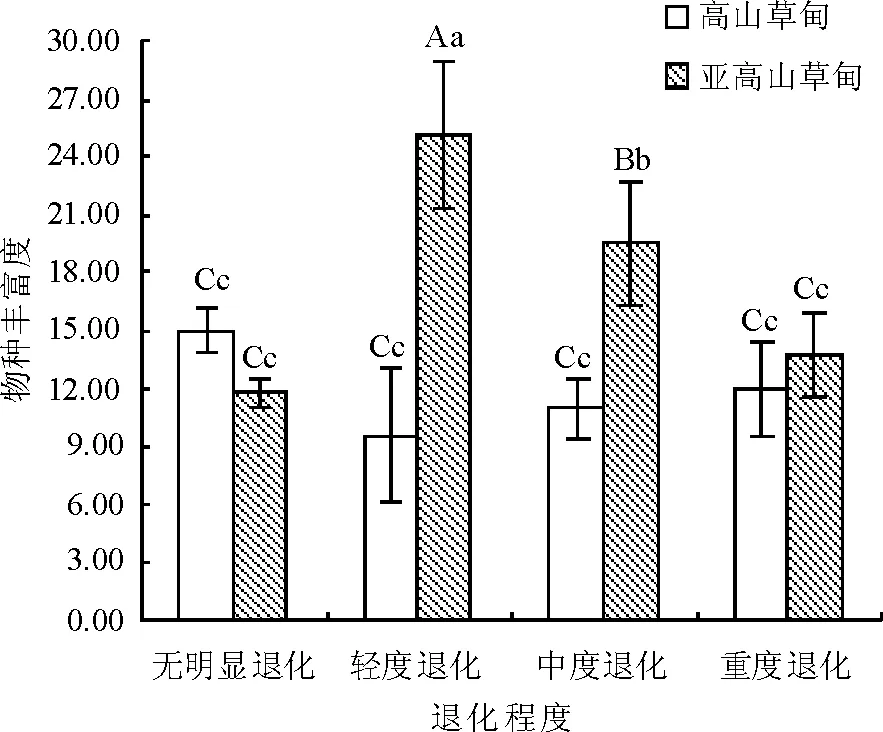
图1 不同退化程度的物种丰富度变化
高山草甸与亚高山草甸退化草地群落优势种存在一定差异,高山嵩草在高山草甸退化草地大量出现,在亚高山草甸退化草地则较少出现;优良牧草老芒麦在高山草甸较少出现;杂类草种类及优势度亦存在差别(表1)。
2.2物种多样性及均匀度变化 高山草甸退化草地物种Shannon-Wiener多样性指数变化不显著,仅无明显退化(2.49±0.02)与中度退化(2.35±0.10)间差异显著(P<0.05);亚高山草甸退化草地物种多样性变化呈“抛物线”趋势,说明放牧干扰对高山草甸多样性影响较小,而对亚高山草甸影响较显著(表2)。
高山草甸均匀度指数(Pielou)变化无规律性,最大值出现在中度退化(0.95±0.03),而轻度退化草地(0.94)与中度退化草地的群落均匀度(0.92)较高,说明植被优势度分配较均匀;亚高山草甸均匀度变化显著,无明显退化到重度退化呈明显上升趋势,说明放牧对高山草甸均匀度影响不及亚高山草甸的显著(表2)。
2.3植被盖度变化 高山草甸与亚高山草甸放牧退化草地盖度均随退化程度增加而下降。高山草甸中,重度退化与无明显退化相比盖度下降了32%,但重度退化盖度仍然达64.00%;亚高山草甸盖度变化梯度不明显,轻度退化(96.00%)与中度退化(83.50%)和重度退化(88.50%)差异极显著(P<0.01)(表2)。
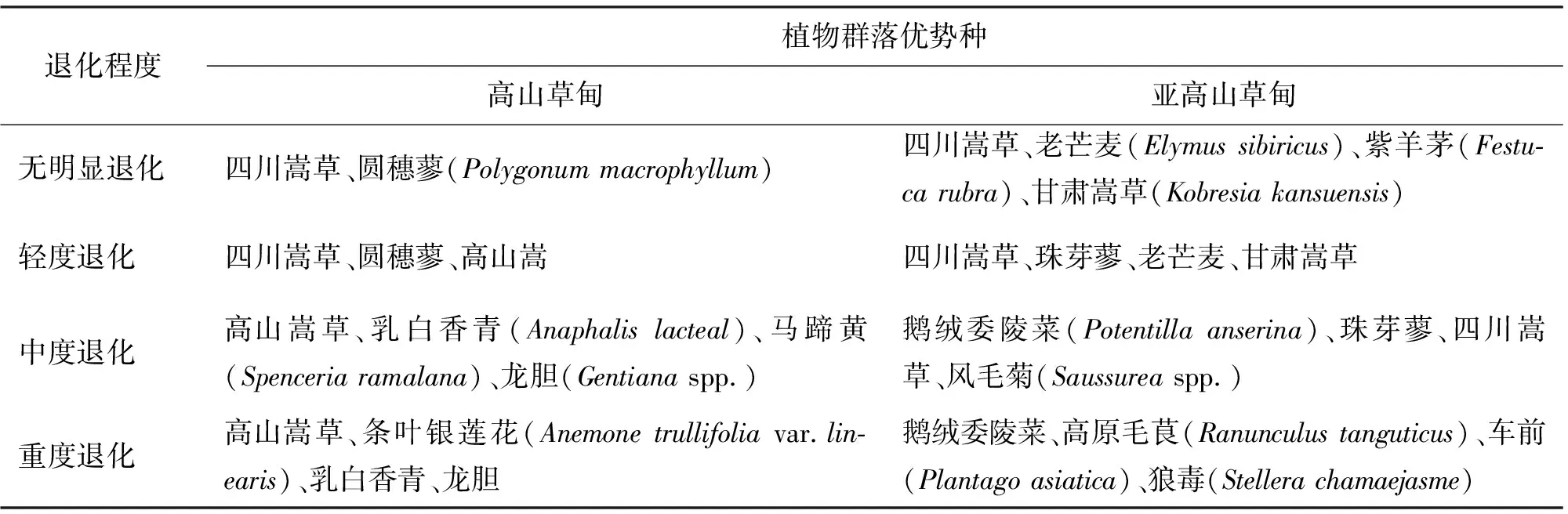
表1 川西北高山草甸与亚高山草甸不同退化程度草地优势种

表2 不同退化程度草地Shannon-Wiener指数、Pielou指数和盖度变化
2.4植被高度变化 高山草甸与亚高山草甸放牧退化草地植被高度随退化程度加剧,呈下降趋势。在无明显退化阶段的最高。高山草甸植被高度无明显退化(14.0 cm)与中度退化(6.0 cm)和重度退化(5.0 cm)差异均极显著(P<0.01),而与轻度退化差异不显著(P>0.05),重度退化较无明显退化下降了72%,处于围栏封育(无明显退化)的草地最高;亚高山草甸差异达极显著(P<0.01),仅中度退化(10.0 cm)与重度退化(4.0 cm)差异不显著(P>0.05),结果均说明放牧干扰降低了草地群落高度。高山草甸高度变化为3.98~13.98 cm,而亚高山草甸为3.26~41.42 cm(图2)。
2.5地上现存量变化 放牧干扰不同程度地使地上现存量减少,除无明显退化草地外,在轻度退化阶段的地上现存量最高。高山草甸退化草地与亚高山草甸退化草地有相同变化规律,但亚高山草甸地上现存量的下降趋势明显高于高山草甸。亚高山草甸重度退化(236.06 g/m2)较无明显退化(1 151.20 g/m2)的降幅达到79%,而处于围栏封育(无明显退化)的草地现存量最高,说明围栏封育有利于现存量的积累,同时亚高山草甸地上现存量均高于高山草甸(图3)。
2.6有毒有害植物地上现存量占群落地上现存量的比例 高山草甸有毒有害植物地上现存量占群落地上现存量的比例呈上升趋势,差异均未达到极显著水平(P>0.01),无明显退化(15.97%)、轻度退化(16.37%)分别与中度退化(27.27%)、重度退化(28.40%)差异显著(P<0.05);亚高山草甸有毒有害植物地上现存量占群落地上现存量的比例变化规律性不明显,无明显退化中比例最小,仅为0.37%,说明围栏对草地优良物种的恢复起到了明显作用,同时抑制了有毒有害植物的生长,降低了地上现存量(表3)。
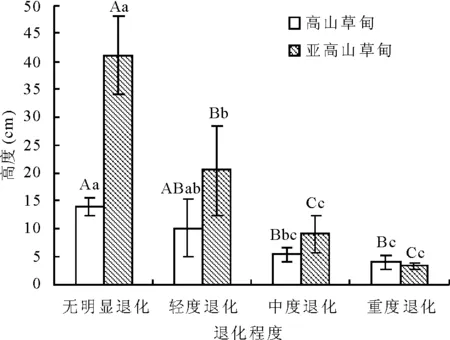
图2 不同退化程度的植被高度变化
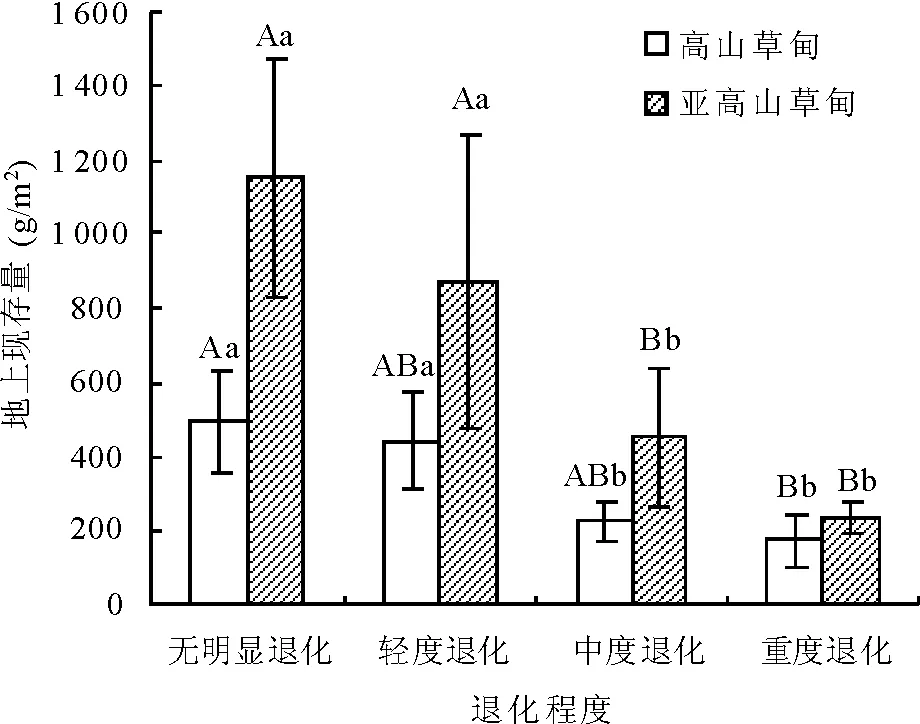
图3 不同退化程度的植被地上现存量变化
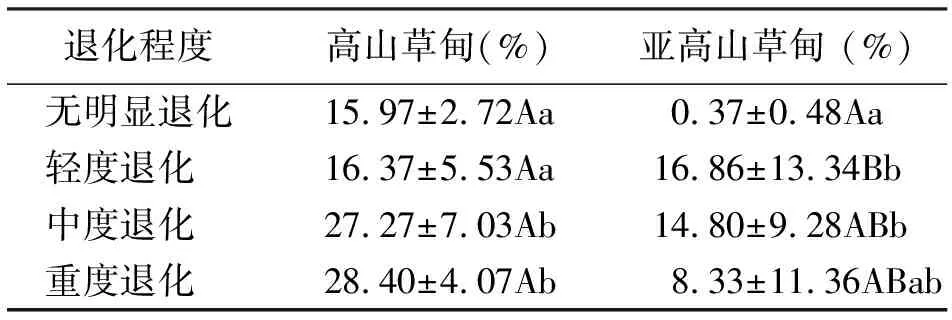
表3 有毒有害植物地上现存量比例变化
3 讨论
在相同退化阶段高山草甸草地的地上生物量和植被盖度均低于亚高山草甸。造成这种差异主要原因是环境条件、植被类型与放牧历史的不同。如亚高山草甸退化草地的植被高度变化极显著的原因是禾本科植物占的比例较大且耐牧性较差。这些差异表明川西北退化草地植被变化特征与其他青藏高原地区的同类研究具有相似性[6-7]。此结果与公延明等[8]对巴音布鲁克高寒草地退化演替阶段植物群落特性研究分析结果相近。
随着退化程度加剧,高山草甸植被盖度呈下降趋势,而亚高山草甸植被盖度的变化规律性差,区分度较低。随着放牧退化程度加剧使群落优势植物种发生更替,耐牧植物增多,而有毒植物条叶银莲花、龙胆、狼毒以及乳浆大戟(Euphorbiaesula)等的大量出现挤占了其他优良牧草生存空间,降低草地质量,物种“小型化”现象明显[9-10]。这种“小型化”现象与杂类草(如:龙胆)等莲座状植物的投影面积较大覆盖地表的特点有关,说明植物种类不同直接影响草地植被盖度的变化,而草地退化对盖度的影响亦由于地区与植被类型差异而发生变化。
亚高山草甸草地的物种丰富度及植被分布均匀度均高于高山草甸草地。在高山草甸草地无明显退化的物种多样性最高则可能与用来计算物种多样性的Shannon-Wienner指数有关,由于该指数为信息指数,以植物种优势度为公式计算的来源,若某些植物种的优势度较高则会使物种多样性指数显著升高。而无明显退化中,由于优势种如四川嵩草、圆穗蓼等植物占据了大量的群落优势度,所以无明显退化的物种多样性指数值均高于其他退化草地;而轻度退化对亚高山草甸具有最高的物种多样性,能促进草地植物群落的更新与稳定,该强度的放牧干扰有利于保持群落稳定性并提高群落生产力[11],与“中度干扰理论”的相关研究结果具有一致性[12]。
高山草甸退化有毒有害植物地上现存量占群落地上现存量比例高于亚高山草甸,其增加趋势是由于家畜对其采食量的下降造成的,这与Lym和Kirby[13]报道乳浆大戟的出现可引起可食草产量下降75%以上而使其所占比例上升的结果一致。
研究区草地生态系统普遍受到人为活动干扰,偏离正常状态[14-15]。对轻度退化草地,建议在合理利用和保护的前提下,维持其生态功能;对中度和重度退化草地,则在条件允许下尽量减少放牧活动干扰,以恢复并提高其生态功能。
4 结论
不同退化程度下的川西北高山草甸与亚高山草甸草地物种丰富度变化呈相反趋势,说明放牧对2类退化草地物种丰富度的影响具有相异性。草地群落优势种存在一定差异,高山嵩草在高山草甸退化草地大量出现而优良牧草老芒麦则较少出现,高山嵩草在亚高山草甸则出现较少。盖度和高度在高山草甸和亚高山草甸上均随放牧强度的增大而下降。高山草甸有毒有害植物地上现存量占群落地上现存量的比例呈上升趋势且比亚高山草甸的比例高。
[1]马玉寿,郎百宁,王启基.“黑土型”退化草地研究工作的回顾与展望[J].草业科学,1999,16(2):5-9.
[2]周寿荣.四川草地资源[M].成都:四川民族出版社,1998:141-153.
[3]邱英,干友民,王钦,等.川西北放牧草地退化分类评价指标体系初探[J].湖北农业科学,2007,46(5):723-726.
[4]任继周.草业科学研究方法[M].北京:中国农业出版社,1998:207-211.
[5]Hutchings M J,de Kroon H.Foraging in plants: the role of morphological plasticity in resource acquisition[J].Advanced Ecology Research,1994,25:159-238.
[6]周华坤,赵新全,周立,等.青藏高原高寒草甸的植被退化与土壤退化特征研究[J].草业学报,2005,14(3):31-40.
[7]汪诗平.青海省“三江源”地区植被退化原因及其保护策略[J].草业学报,2003,12(6):1-9.
[8]公延明,胡玉坤,阿德力·麦地,等.巴音布鲁克高寒草地退化演替阶段植物群落特性研究[J].干旱区资源与环境,2010,24(6):149-152.
[9]干友民,李志丹,王钦,等.川西北亚高山草甸放牧退化演替研究[J].草地学报,2005,13(S1):48-52.
[10]韩友吉,陈桂琛,周国英,等.青海湖地区高寒草原植物个体特征对放牧的响应[J].中国科学院研究生院学报,2006,23(1):118-122.
[11]王德利,祝廷成.不同种群密度状态下羊草地上部生态场、生态势、场梯度及其季节性变化规律研究[J].生态学报,1996,16(2):121-127.
[12]宝音陶格涛,刘美玲,李晓兰.退化羊草草原在浅耕翻处理后植物群落演替动态研究[J].植物生态学报,2003,27(2):270-277.
[13]Lym R G,Kirby D R.Cattle foraging behavior in leafy spurge (Euphorbiaesula)-infested rangeland[J].Weed Technology,1987,1:314-318.
[14]王岩春,干友民,费道平,等.川西北退牧还草工程区围栏草地植被恢复效果的研究[J].草业科学,2008,25(10):18-19.
[15]干友民,罗元佳,周家福,等.川西北沙化草地生态恢复工程对沙地植被群落的影响[J].草业科学,2009,26(6):51-56.
猜你喜欢
戏曲研究(2022年2期)2022-10-24 01:54:12
黑龙江工程学院学报(2020年5期)2020-10-21 05:37:10
水土保持研究(2019年6期)2019-10-19 03:33:32
绿色科技(2017年6期)2017-04-20 08:13:04
河北林业科技(2016年5期)2016-11-08 03:13:30
西南农业学报(2016年5期)2016-05-17 05:42:44
西藏大学学报(自然科学版)(2015年1期)2015-12-25 11:43:44
防护林科技(2015年5期)2015-06-10 11:42:52
海南热带海洋学院学报(2015年5期)2015-03-14 09:51:07
河南科技(2014年16期)2014-02-27 14:13:25
