
不同利用方式下新疆春秋牧场植物群落特征变化趋势
2011-04-25 09:40靳瑰丽阿德列提邓新疆再努然木阿不都艾尼
草业科学 2011年7期
闫 凯,靳瑰丽,刘 伟,阿德列提,邓新疆,再努然木·阿不都艾尼
(1.新疆草原总站,新疆 乌鲁木齐 830049;2.新疆农业大学草业与环境科学学院 新疆草地资源与生态实验室,新疆 乌鲁木齐 830052;3.新疆新源县畜牧局,新疆 新源 835800)
春秋牧场在一年中分春、秋两季利用,利用时间长达半年之久,在畜牧业生产发展与生态环境保护建设中有着极其重要的作用,但由于其分布格局、利用时间上的特殊性决定了春秋牧场在利用上存在生态系统脆弱、极易受损,却仍然担负着极大放牧压力的特点,再加上人为破坏及保护意识低下,从而加剧了草地的退化且成为退化最为普遍、严重的地段[1-3]。构成草地的主体——植物,其生长具有季节间和年度间的动态性,在自然和人为不断干预和影响下,发生着不同程度的演替[4-5]。利用方式的差异性能够给草地带来不同程度的影响,导致植物个体大小变化,相应种群生态位也发生改变,因而必然引起群落在总体结构上的重新组织[6],利用方式合理可以改良草地,有助于土壤碳库的稳定[7],利用方式不当使草地退化。因此,合理的利用方式对草地的健康发展具有重要的意义。本研究以此为切入点,在对不同利用方式下春秋牧场植物群落结构组成、高度、盖度、密度、地上生物量进行测定分析的基础上,研究不同利用方式下植物群落特征变化趋势,为草地合理利用方案的制定提供科学依据。
1 材料与方法
1.1研究区概况 项目实施地点设立在新源县则克台镇阿西勒村春秋牧场,43°32′49″~43°33′58″ N,83°01′23″~83°14′24″ E,海拔856~907 m。属于平原河谷地区,为干旱区。年均气温在3~5 ℃,无霜期167 d左右,年降水量100~160 mm,≥0 ℃年积温为3 511 ℃·d,≥10 ℃年积温3 081 ℃·d。
该牧场为白羊草(Bothriochloaischaemum)+杂类草草原,春季有短生、类短生植物发育,夏季一年生草本发育显著,秋季木地肤(Kochiascoparia)半灌木长势良好,在年降水量和利用强度适宜时,夏季可打草1次。由于放牧强度较大,牧民保护意识低下,加之缺乏科学的管护措施,该类草地退化普遍。
目前阿西勒村该类草地已承包到户,于2007年进行了围栏划分,分属67户牧民,独立经营。通过对牧户的调查与分析得知,不同牧户对该类草地有4种利用方式,由于牧民放牧利用方式及利用强度不同,草地退化程度也存在一定的差异。
1)打草型:不放牧,仅夏季打草1次;2)自由放牧型:春秋季连续自由放牧;3)分段放牧型:将草地围栏划分为两段,50%春秋季自由放牧,50%秋季自由放牧+夏季打草1次;4)划区轮牧型:春秋季四区轮牧,轮牧间隔6~10 d。
1.2取样方法 分别对研究区21户牧民生产方式及春秋牧场利用现状进行入户调查,初步了解牧户生产经营现状及草地发展概况。筛选放牧畜种、放牧强度相对一致的牧户研究不同利用方式对草地带来的影响,其中打草型2户,自由放牧型2户,分段放牧型3户,划区轮牧型2户,进行春、夏、秋季草地群落跟踪调查。典型牧户春秋牧场面积和家畜数量不等,但放牧强度约16羊单位/(hm2·时段),放牧家畜自然头数羊∶牛∶马为15∶2∶1左右。
于2009年4月中旬、6月中旬、8月下旬,即草地春季放牧前、夏季打草前和秋季放牧前3个时段,以各牧户草地为大样地,在其典型地段布置5个1 m×1 m样方,各季节测定样方布置地点不重复。
1.3测定指标及方法 在各样方内,记录植物种类, 分种测定其高度、盖度、密度、地上生物量(鲜草),并在样地内测定频度,并计算植物重要值[8]。
式中,RC为草地植物的相对盖度,RB为草地植物的相对地上鲜草产量,RD为草地植物的相对密度,RF为草地植物的相对频度。
1.4数据处理 测得的数据用Excel 2000进行统计,运用SPSS软件对数据进行处理,采取One-way ANOVA和LSD对数据进行方差分析和显著性分析。
2 结果
2.1不同利用方式下草地植物群落结构变化 植物群落的物种组成是决定草地植物群落性质、结构和功能的重要指标,而环境因子对其具有决定性作用,放牧等人为活动会对其产生明显的影响[9]。
分别选取各样地重要值排在前3位的植物分析群落组成(表1)。结果表明,4种利用方式从春季到秋季,植物群落结构发生了明显的变化,物种数从春季到秋季逐渐减少,春季禾草和短生、类短生植物发育明显,夏季以一年生禾草和杂类草为主,而秋季以木地肤、白羊草和苔草等多年生植物为主。但不同利用方式下草地群落物种组成具有一定的差异性。
4月群落物种数以划区轮牧最多,达30种,划区轮牧是其他3种利用方式的1.5倍左右,其他3种利用方式间相差不大(表1)。可见,划区轮牧可以提高春季草地物种丰富度,更有益于草地多样性的维持。在群落组成上,自由放牧、分段放牧和划区轮牧3种利用方式均为禾草+短生、类短生草本,而打草利用方式为短生、类短生草本+禾草,可见放牧更能刺激翌年禾本科牧草的分蘖,而割草更能促进杂类草的生长。
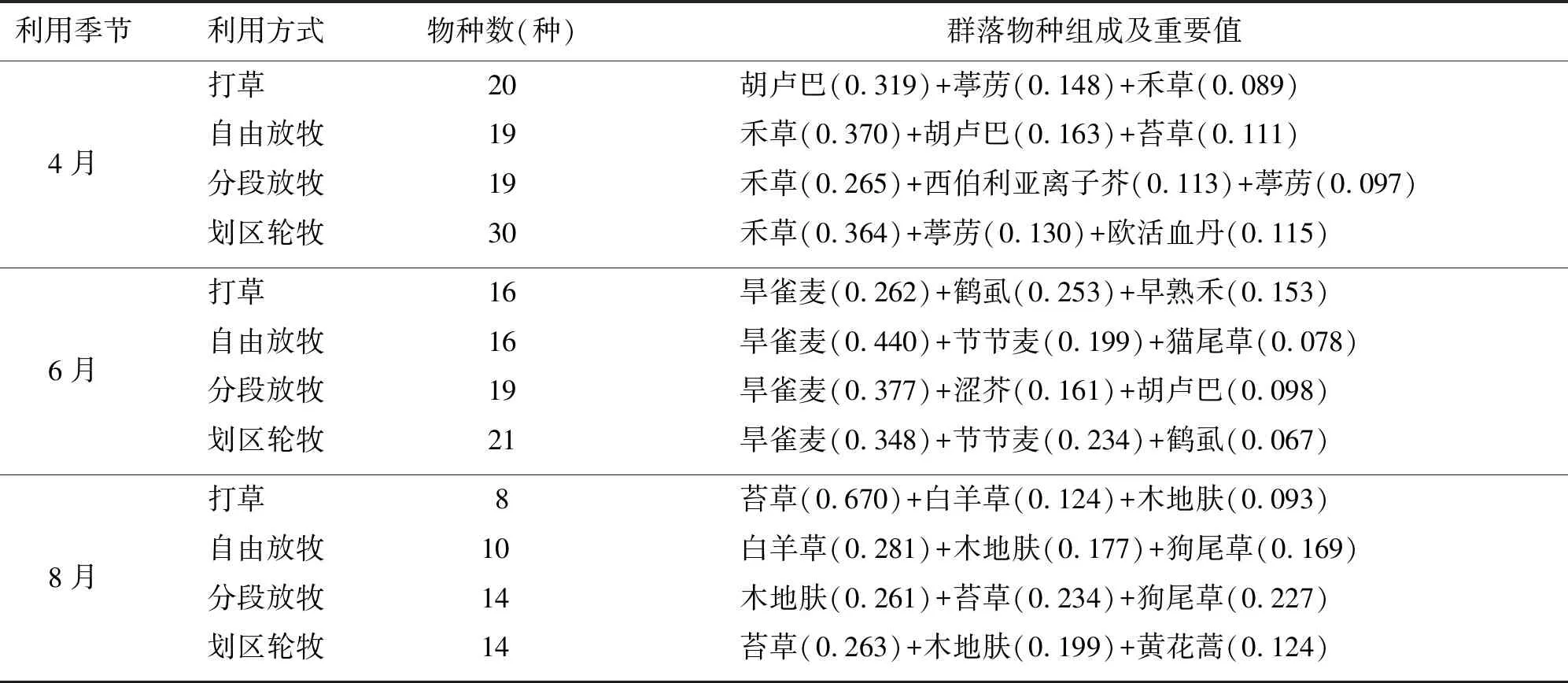
表1 不同利用方式草地植物组成变化
6月群落物种数仍以划区轮牧最多,而分段放牧次之,打草和自由放牧相当,可见有计划的放牧和划区轮牧利于草地植物在生长最旺盛的季节进行物种多样性的维持。在群落组成上,自由放牧以旱雀麦+节节麦+猫尾草为主,禾本科牧草优势地位突出;其他3种利用方式大多为一年生禾草+杂类草,但打草地为旱雀麦+鹤虱+早熟禾,且旱雀麦与鹤虱重要值相差不大;分段放牧草地为旱雀麦+涩芥+胡卢巴,由于早春50%草地不放牧,使短生、类短生植物发育良好;划区轮牧草地为旱雀麦+节节麦+鹤虱,禾草优势地位仅次于自由放牧(表1)。进一步说明了放牧比打草更能促进禾草的生长,而对杂类草有抑制作用。
8月,群落物种数以打草最少,降低至8种,而分段放牧和划区轮牧均为14种,是打草的1.75倍,自由放牧居中,为10种,可见放牧干扰下草地植物秋季物种丰富度较打草高,并且分段放牧对植物物种多样性维护的潜在效能能发挥出来。在群落组成上,虽然苔草、白羊草、木地肤等多年生植物作用凸显,但各利用方式之间无明显的规律性变化,相比之下,自由放牧草地虽然为白羊草+木地肤+狗尾草,但多年生植物的优势地位略逊与其他利用方式(表1)。长期自由放牧会不利于草地植物群落结构的稳定,具有潜在风险。
2.2不同利用方式下草地植物群落高度变化 草地群落的高度是表示一个群落的数量特征,反映群落经济性状大小的指标。4种利用方式群落高度均表现为6月>8月>4月(图1),即春季转场后,草地经过近2个月的再生生长,夏季植物生长旺季草地群落达到年度内的最高值,开始打草。秋季随着枯黄期的到来,相对于6月,各利用方式下草地植物群落高度下降明显。不同利用方式对草地群落高度产生一定的影响。
4月,草地群落高度表现为打草>分段放牧>自由放牧>划区轮牧,打草型显著高于其他放牧利用方式(P<0.05),是其他利用方式2倍多;自由放牧、分段放牧、划区轮牧3种放牧利用方式间差异不显著(P>0.05)。6月,草地群落高度表现为划区轮牧>打草>自由放牧>分段放牧,划区轮牧草地群落高度高达47.74 cm,且与分段放牧差异显著(P<0.05)。8月草地群落高度为划区轮牧>分段放牧>自由放牧>打草,划区轮牧显著高于打草型(P<0.05)。可见,自由放牧、分段放牧、划区轮牧3种放牧利用方式下,践踏和采食影响了翌年春季草群高度,均低于打草利用方式;然而从高度增长量来看,自由放牧、分段放牧、划区轮牧3种放牧利用方式均大于打草方式,并最终高于打草型,其中划区轮牧更能增加草地休养生息的机会,有利于草群高度的提高,在6月和8月均为最高值。

图1 不同利用方式草地植物群落高度变化
2.3不同利用方式下草地植物群落盖度变化 草地植被物种盖度变化可以反映该草地的类型、植物同化面积的大小、生物量积累的多少[10]。4种利用方式群落盖度除打草为4月>6月>8月,其余3种放牧方式均为6月>4月>8月(图2)。打草地春季不放牧,加之短生、类短生植物大量发生,使得其春季盖度与夏季相差不大,自由放牧、分段放牧、划区轮牧3种放牧方式春季放牧转场后,草地经过近2个月的再生生长,夏季植物生长旺季草地群落达到年度内的最高值,开始打草。秋季随着枯黄期的到来,相对于6月份,各利用方式下草地植物群落盖度下降明显,达到年内最小值。不同利用方式间群落盖度的变化趋势与群落高度基本一致。
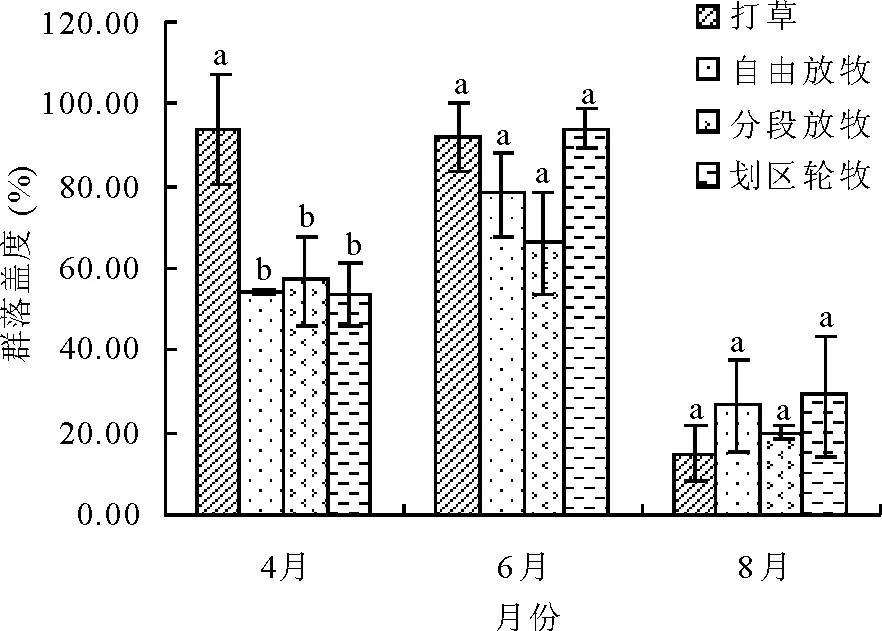
图2 不同利用方式草地植物群落盖度变化
4月植物生长初期,草地群落盖度为打草>分段放牧>自由放牧>划区轮牧,打草型草地由于春季不放牧,植物群落盖度达到93.70%,显著高于其他3种利用方式(P<0.05),其他3种放牧利用方式间差异不显著(P>0.05),其中分段放牧由于早春有一半草地不放牧,所以群落平均盖度略高于自由放牧和划区轮牧草地。6月植物生长迅速,草地群落盖度表现为划区轮牧>打草>自由放牧>分段放牧,但差异不显著(P>0.05)。8月草地群落盖度为划区轮牧>自由放牧>分段放牧>打草,但差异不显著(P>0.05)。可见,打草和分段放牧春季不放牧或少放牧仅提高春季群落盖度,但划区轮牧更能从物种多样性和再生能力上提高群落盖度,使其在夏季和秋季具有较高的群落盖度。
2.4不同利用方式下草地植物群落密度变化 密度是草地植物重要的数量特征,其多少在一定程度上可反映草地多度的大小。由于从春季到秋季物种数不断降低,加之春季早春植物大量爆发,夏季一年生植物的繁殖,使得4种利用方式草地群落密度基本上表现为4月>6月>8月(图3)。各种利用方式下,密度变化存在一定的差异。
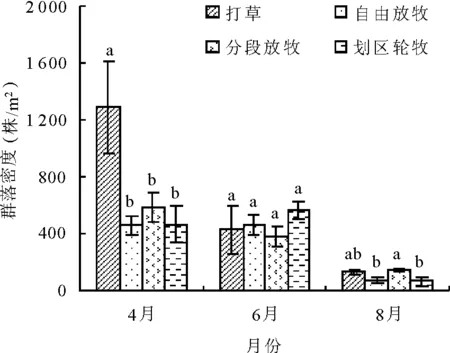
图3 不同利用方式草地植物群落密度变化
4月草地群落密度为打草>分段放牧>划区轮牧>自由放牧,打草型高达1 289 株/m2,显著高于其他放牧利用类型(P<0.05),而3种利用方式间差异不显著。春季打草型完全不放牧,短生类短生植物大量爆发,使得其群落密度最高,而分段放牧地50%春季不放牧,植物群落密度略高于其他2种放牧类型。6月,草地群落密度为划区轮牧>自由放牧>打草>分段放牧,但各利用方式间差异不显著。划区轮牧型由于物种数量相对较多使得群落密度有所提高。8月,草地群落密度为分段放牧>打草>自由放牧>划区轮牧,分段放牧显著高于自由放牧和划区轮牧(P<0.05)。受季节变化、物种变更等多重因素的影响,各利用方式的群落密度在季节间排列顺序变动幅度较大,春季打草型最大,夏季划区轮牧型最大,而秋季分段放牧型最大,其原因有待于进一步研究。
2.5不同利用方式下草地植物群落地上生物量变化 地上生物量是草地初级生产力的直观体现,不同利用方式会使其发生不同程度的变化。各利用方式草地群落地上生物量均为6月>4月>8月(图4),夏季达到最高值,打完草后,由于干旱少雨,牧草再生能力不高,因此秋季生物量较低。
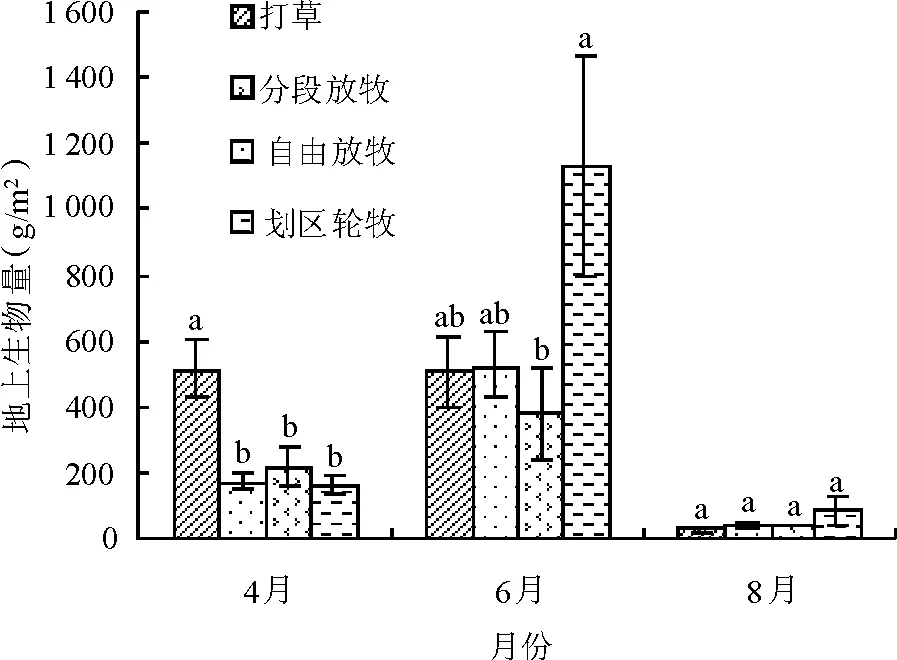
图4 不同利用方式草地植物群落地上生物量变化
草地群落地上生物量,4月表现为打草>分段放牧>自由放牧>划区轮牧,打草型显著高于自由放牧、分段放牧、划区轮牧3种放牧利用型(P<0.05),3种放牧利用方式之间差异不显著(P>0.05);6月表现为划区轮牧>自由放牧>打草>分段放牧,划区轮牧型生物量提高到1 128 g/m2,显著高于分段放牧型(P<0.05);8月表现为划区轮牧>自由放牧>分段放牧>打草,各利用方式间差异不显著(P>0.05)。暖季累计鲜草产量为划区轮牧>打草>自由放牧>分段放牧,分别为1 366.21、1 049.84、737.08和631.20 g/m2,划区轮牧促进了草地地上生物量的积累。
3 讨论与结论
很多研究表明, 与自由放牧相比, 轮牧更有利于群落地上生物量的恢复和提高[11],随着轮牧年限的延长,划区轮牧区的地上生物量显著高于自由放牧区,因此,轮牧有利于群落生物量的积累[12]。本研究轮牧草地暖季累计鲜草产量最高,是自由放牧的近2倍。并且本研究表明在物种数上,划区轮牧型在各季节物种数均最大,其更有益于草地多样性的维护。
本研究群落各项数量特征为春季打草型显著高于其他放牧利用类型,而从夏季开始,打草型逐渐低于其他利用方式,到秋季成为4种利用方式中的最低者,取而代之的是划区轮牧在夏季和秋季各项数量特征升值最高;并且在群落组成上,3种放牧类型更能刺激翌年禾本科牧草的分蘖,而打草型则促进杂类草的生长。在半干旱热带稀树草原和半天然草地中的研究结果表明,割草处理的草地由于连续割草会带走地上生物量且不返还动物排泄物,如果割草量过多就会导致土壤养分流失[13-14];对天然羊草(Leymuschinensis)草原的研究结果表明,割草能引起土壤贫瘠化和密度增大,刈割的时间越长,对种子生产的影响就越严重,长期连年刈割的小区穗序发育不良,结实的籽粒瘦秕[15]。因此长期打草不放牧,使草地的经济和生态效益逐渐丧失。
分段放牧是当地中小生产规模牧户使用的一种放牧方式。薛睿等[16]以羊为单位设计载畜率水平对内蒙古典型草原群落研究表明,放牧和割草轮换的混合利用方式下,群落地上净初级生产力显著高于传统放牧和传统割草方式,可以减缓放牧带来的持续性压力,对草地具有一定的保护作用,特别是在过度放牧的情况下,混合利用方式尤佳。因此,本研究中分段放牧以自然生产状态羊∶牛∶马为15∶2∶1放牧畜种,放牧强度约16羊单位/(hm2·时段)为基础,虽然草地群落各项数量特征并不占绝对优势,但在维护草地群落物种上比自由放牧和打草型较高,因此对于中小生产规模牧户适当调节畜群结构和放牧压力,分段放牧不失为一种经济的放牧管理方式,而且50%早春不放牧草地需要在两块地之间轮换,可提高群落生产和生态性能。
综合对比各利用方式下白羊草+杂类草草原植物群落动态变化趋势,划区轮牧无论在维护草地群落结构稳定性,还是在提高群落数量特征和生产性能上均具有明显的优势;打草不放牧不利于草地的更新和繁衍;长期自由放牧则不利于草地可持续发展;而分段放牧兼顾了轮牧和打草的利用方法,合理调节放牧畜群和放牧压力,将会在提高草地生产生态效益上具有潜在的优势。
放牧是一种高度复杂的干扰方式,是天然草地的普遍利用方式,对植物群落既有积极作用,也有消极作用,其作用效果受多种因素的影响,具有滞后性[17-19]。本研究是在维持不同利用方式3年的基础上进行的,所以应详细调查与记录草地本底数据,进行多年长期动态监测,进一步丰富研究结果。
[1]许鹏.新疆草地资源及其利用[M].乌鲁木齐:新疆科技卫生出版社,1993.
[2]靳瑰丽,朱进忠,陈乐宁,等.伊犁绢蒿荒漠受损草地植物群落春季演变趋势分析[J].草业科学,2007,24(10):26-29.
[3]杨智明,李建龙,杜广明,等.宁夏滩羊放牧系统草地利用率及草畜平衡性研究[J].草业学报,2010,19(1):35-41.
[4]赵万羽,李学森,阿依丁.天山北坡山地割草场改造利用技术研究[J].草食家畜,2000(增刊):38-42.
[5]韩文军,春亮,侯向阳.过度放牧对羊草杂类草群落种的构成和现存生物量的影响[J].草业科学,2009,26(9):195-199.
[6]王炜,梁存柱,刘钟龄,等.草原群落退化与恢复演替中的植物个体行为分析[J].植物生态学报,2000,24(3):268-274.
[7]孔玉华,姚风军,鹏爽,等.不同利用方式下草地土壤碳积累及汇/源功能转换特征研究[J].草业科学,2010,27(4):40-45.
[8]任继周.草业科学研究方法[M].北京:中国农业出版社,1998.
[9]闫玉春,唐海萍.围栏禁牧对内蒙古典型草原群落特征的影响[J].西北植物学报,2007,27(6):1225-1232.
[10]魏斌.生态环境功能评价可持续发展之路[M].北京:北京大学出版社,1994.
[11]朱桂林,卫智军,杨静,等.放牧制度对短花针茅群落植物种群地上生物量的影响[J].中国草地,2002,24(3):15-19.
[12]邢旗,双全,金玉,等.草甸草原不同放牧制度群落物质动态及植物补偿性生长研究[J].中国草地,2004,26(5):26-31.
[13]Augustine D,McNaughton S J.Interactive effects of ungulate herbivores, soil fertility and variable rainfall on ecosystem processes in a semi-arid savanna[J].Ecosystems,2006(9):1242-1256.
[14]Ilmarinen K,Mikola J,Nissinen K,etal,Role of soil organisms in the maintenance of species-rich semi-natural grasslands through mowing[J].Restoration Ecology,2009(17):78-88.
[15]杨允菲.天然羊草草地在放牧和刈割条件下的种子生产性能[J].中国草业科学,1988,5(6):30-33.
[16]薛睿,郑淑霞,白永飞.不同利用方式和载畜率对内蒙古典型草原群落初级生产力和植物补偿性生长的影响[J].生物多样性,2010,18(3): 300-311.
[17]Savory A,Parsons L D.The Savory grazing method[J].Rangelands,1980(2):234-237.
[18]Heady H F,Pitt M D.Seasonal versus continuous grazing on annual vegetation of Nothern California[J].Rangelands,1979(6):231-232.
[19]Hoveland C S,McCann M A,Hill N S.Rotational vs.continuous stocking of beef cows and calves on mixed endophyte free tall fescue bermudagrass pasture[J].Journal of Production Agriculture,1997,10(2):245-250.
猜你喜欢
数学物理学报(2021年4期)2021-08-30
今日农业(2020年19期)2020-12-14
黑龙江工程学院学报(2020年5期)2020-10-21
水土保持研究(2019年6期)2019-10-19
小学生学习指导(低年级)(2018年11期)2018-12-03
农民致富之友(2018年20期)2018-11-25
中国农业科学(2017年8期)2017-05-03
甘肃畜牧兽医(2016年12期)2016-09-21
太空探索(2016年9期)2016-07-12
海南热带海洋学院学报(2015年5期)2015-03-14
