
三种盐生植物在沿海滩涂湿地土壤中微生物数量的变化※
2011-04-16 02:56:46李洪山申玉香
农业现代化研究 2011年2期
李洪山,申玉香
(江苏盐城工学院化学与生物工程学院,江苏盐城224051)
我国海岸线漫长,北起辽宁鸭绿江口,南至广西北仑河口,沿海滩涂分布十分广泛,滩涂总面积约200万hm2,并且沿海滩涂在泥沙来源丰富的海岸带仍在淤长,全国沿海滩涂每年约淤长300 km2,江苏省沿海滩涂面积据沿海省市之首,面积达全国滩涂总面积的1/4左右,江苏沿海滩涂生态类型多样,开发好滩涂资源对加快经济特别是沿海经济带的发展有着举足轻重的作用[1,2]。
滩涂土壤资源科学利用是滩涂经济发展的基础,土壤微生物数量及功能是与土壤碳、氮等元素的循环过程和土壤养分的矿化过程关系密切,对有机物质的分解转化、养分的转化和供应起着重要的主导作用。不同植物群落生境中土壤微生物区系有差异,盘锦芦苇湿地土壤微生物不同时间动态上是细菌数量最大,其次是放线菌,最少的是真菌。在0cm~10cm、10cm~20cm和20cm~30cm三个层次中以中层的细菌、放线菌和真菌数量最大。细菌、放线菌数量都是从6月的最高值开始减少,到8月达到最低,而后逐渐增加;真菌数量则表现为从6月开始逐渐增加,至9月达到最大值[3]。江苏省滨海县滩涂盐碱地土壤微生物区系与对照菜园土壤相比,除细菌数量与对照无显著差异外,真菌、放线菌、解磷菌及钾细菌数量均比对照土壤明显降低[4]。在江苏盐城新洋港互花米草盐沼地,细菌在所有的微生物组成中最为丰富。数量总体表现为细菌>放线菌>真菌,分布表现为米草带>碱蓬带>光滩带。真菌在数量上明显低于细菌2~3个数量级,分布表现为米草带≥碱蓬带>光滩带。放线菌在数量上总体表现出碱蓬带>米草带>光滩带[5]。滩涂盐生植物资源丰富,一些植物群落种类如碱蓬、芦苇等在滩涂土壤改良中作用显著[6]。植物根系给土壤微生物提供了一个特殊的生态环境,在滩涂植物群落不断向耕地植物群落演替的过程中,土壤性质的变化成了群落演替的主要动力。由此研究盐土植物群落与土壤微生物特征的变化同一性,是合理经营和利用一切自然资源的理论基础,有助于对自然生态系统和人工生态系统进行有效的控制和管理,并且可指导退化生态系统恢复和重建,其与农、林、牧和人类经济活动紧密相连。
1 材料与方法
1.1 土壤样品采集
土壤样品采集分别于2008年6月(夏季)和12月(冬季)进行。所选样地在江苏省东台市沿海现代农场境内(32°86'N,120°70'E),在农场境内选择盐地碱蓬群落、互花米草群落和芦苇群落三种不同生境土壤,铲除表层2cm土壤,采集2cm-10cm土层土壤作为实验样品,每个样品采用对角线五点取样,混合后装入塑料自封袋备用。同时在盐城市农业科学院实验农场水稻田(33°38'N,120°32'E)(以下简称水稻田)采样2cm-10 cm土层土壤作为对照样品。
1.2 土壤理化性质分析
土壤有机质含量采用硫酸-重铬酸钾氧化法(水合热法)[7];全氮采用半微量凯氏法[7];速效磷采用酸溶-钼蓝比色法[7];水溶性盐测定采用残渣烘干法[7]。用酸度计直接测定土壤溶液pH值(溶液为干土壤与去离子水的质量比为1:5)。
1.3 土壤微生物的分离和计数
无菌环境中称取10g土样于装90m l无菌水的三角瓶中,三角瓶内放置少量无菌玻璃珠,震荡15min使土水混合均匀。用稀释平板法,每个样品做三个重复。在预备性实验的基础上,选择合适稀释度,分离培养微生物。细菌采用牛肉膏蛋白胨琼脂培养基,放线菌采用高氏一号琼脂培养基,真菌采用添加孟加拉红和链霉素的马丁氏培养基,固氮菌采用阿须贝无氮培养基,纤维素分解菌用羧甲基纤维素钠(CMC-Na)培养基,接种后在25℃~28℃下培养,细菌、放线菌和真菌分别在接种后第3d、第7d和第5d进行计数[8,9]。每个土样重复3次培养,以平均数分析。
1.4 数据统计分析方法
数据以烘干土壤重计算,用各重复间的平均数作为统计数。采用DPS2.0统计分析软件对数据进行F测验和t测验,比较不同采样时间之间、不同土壤样品之间的差异性。
2 结果与分析
2.1 不同植物群落生境土壤主要理化性质分析
盐生植物生长在滩涂生态系统中,滩涂含盐分较高,滩涂不同植物对盐分的抗耐受能力有差异,同时,滩涂植物生长对土壤性质变化也有作用。结果表明,农科院稻田土壤有机质含量明显大于滩涂植物生境土壤,在生长季氮素丰富,与土壤营养相关的指标如全氮、速效磷等也高于滩涂盐生植物群落生境土壤,其中以全氮指标值差异更为显著。芦苇对水分的适应幅度很宽,对土壤盐分要求不严格,从土壤湿润到长年积水,从水深几厘米至1m以上,都能形成芦苇群落,每到秋冬季节芦苇枯黄,茎叶落地,土壤中植物纤维丰富。互花米草常年生长在岸边的流水和湿地中,土壤中生物丰富,米草生境土壤中有机物来源多,含量居三种盐生植物生境之首。与农科院稻田土壤相比,滩涂生境盐分含量较高。

表1 不同植物群落生境土壤主要理化性质
2.2 不同植物群落生境土壤细菌数量变化
相对而言,细菌是土壤微生物中数量最多的一个类群,但在不同类型土壤中的数量有差异。研究表明,农科院水稻田夏季土样中细菌数量达到(420±20.07)×106/g干土样,冬季土样中数量为(315±39.68)×106/g干土样,都比同期其他植物生境内土样中细菌数量明显增多,分别是夏季米草生境土样的16.58倍、碱蓬的23.33倍、芦苇的32.31倍,秋季米草生境土样的18.18倍、碱蓬的24.23倍和芦苇的45倍。取样时间不同,细菌数量也有差异,总的趋势是夏季的多于冬季的。在农科院稻田中夏季细菌数量在α=0.01水平上显著多于冬季的(t=4.09**)。三种盐生植物生境土样中的细菌在数量上都是夏季的高于冬季的,但在α=0.05水平上都没有差异。植物生境不同,数量差异性也不同(图1)。同一季节,不同植物生境土样之间细菌数量在p=0.01水平上有差异性(F(夏季)=5.65**,F(冬季)=9.20**)。细菌数量在米草生境土壤中最多,其次是盐地碱蓬生境的,最后是芦苇生境的。夏季米草生境细菌数量与芦苇生境细菌数量在p=0.05水平上有显著差异性,碱蓬生境细菌数量与互花米草和芦苇生境细菌数量都未达显著性差异。冬季的米草生境土样和碱蓬生境土样之间在细菌数量上没有显著性差异,两者与芦苇生境细菌数量之间都有显著性差异。碱蓬生境土样的细菌数量与互花米草和芦苇生境细菌数量之间在p=0.01水平上没有显著性差异,但米草生境细菌数量与芦苇生境细菌数量之间在p=0.01水平上有极显著差异。

图1 不同盐生植物生境中土壤细菌数量差异
2.3 不同植物群落生境中土壤真菌数量变化
不同植物群落生境中土壤真菌数量差异性不显著,结果表明,夏季土样的土壤真菌数量幅度在15×106/g干土样~23×106/g干土样,以农科院水稻田土壤中数量最少,为(15±3)×106/g干土样;冬季土样中的土壤真菌数量幅度在1.33×106/g干土样~5×106/g干土样,以农科院水稻田土壤中偏多(5±1)×106/g干土样。不同盐生植物生境土壤中的真菌数量相仿,相互之间没有显著性差异,夏季、冬季的趋势一致(F值分别为0.65、0.64)。土壤真菌数量有明显的季节性差异,夏季时的数量值与冬季的土壤真菌数量在α=0.01水平上有极显著性差异(图2)。

图2 不同盐生植物生境中土壤真菌数量差异
2.4 不同植物群落生境中土壤放线菌数量变化
放线菌与土壤腐殖质的含量有关,对土壤中的物质转化也具有一定的作用。农科院稻田土壤中的放线菌数量夏季为(29±4.58)×106/g干土样,冬季为(26.33±5.51)×106/g干土样,与三种盐生植物生境土壤中的数量相近,在同一季节,不同盐生植物土壤之间的放线菌数量在p=0.05水平上有差异(F(夏季)=6.66**,F(冬季)=11.8*),在互花米草生境土壤中放线菌数量偏多,在夏季,互花米草生境中土壤放线菌数量与芦苇生境中放线菌数量之间有显著性差异。在冬季,互花米草生境土壤中放线菌数量与盐地碱蓬和芦苇生境土壤中放线菌数量之间都有显著性差异。三种盐生植物生境土壤中放线菌数量在夏季与冬季之间均未达α=0.05水平上的差异,但在数值上是夏季的大于冬季的(图3)。

图3 不同盐生植物生境中土壤放线菌数量差异
2.5 不同植物群落生境中土壤纤维素分解菌数量变化
纤维素分解菌数量以冬季土壤中的大于夏季土壤中的,农科院水稻田土壤中纤维素分解菌数量夏季的为(403±18.33)×104/g干土样,冬季的为(484±19.97)×104/g干土样,均明显多于其他三种盐生植物生境中的土壤纤维素分解菌。在盐生植物生境中,夏季土壤中纤维素分解菌数量以芦苇的最多,盐地碱蓬土壤中纤维素分解菌数量次之,互花米草土壤中纤维素分解菌数量最少,三种盐生植物土壤中纤维素分解菌数量相互之间均在p=0.01水平上有差异性。冬季土壤的趋势与夏季的一致(F(夏季)=143.7**,F(冬季)=112*)。在不同季节相同土壤生境中,纤维素分解菌数量在α=0.05水平上没有差异性。

图4 不同盐生植物生境中土壤纤维素分解菌数量差异
2.6 不同植物群落生境中土壤固氮菌数量变化
在农科院水稻田土壤中固氮菌数量以冬季的高于夏季的,分别为(112±17.06)×104/g干土样,(78±10)×104/g干土样,而在三种盐生植物生境土壤中均以夏季的高于冬季的,且除芦苇生境土壤固氮菌外,其余土壤生境的固氮菌数量在季节之间均在α=0.01水平上有差异性。以夏季互花米草生境土壤中固氮菌数量最多,且与其它两种生境土壤中固氮菌在p=0.05水平上差异显著。在冬季,三种盐生植物生境土壤中固氮菌数量之间没有差异性(F(夏季)=9.5*,F(冬季)=0.6)。固氮菌可以增加土壤氮素,其数量与土壤中氮素含量有关联,土壤中氮源丰富,则可能对固氮菌有抑制作用。
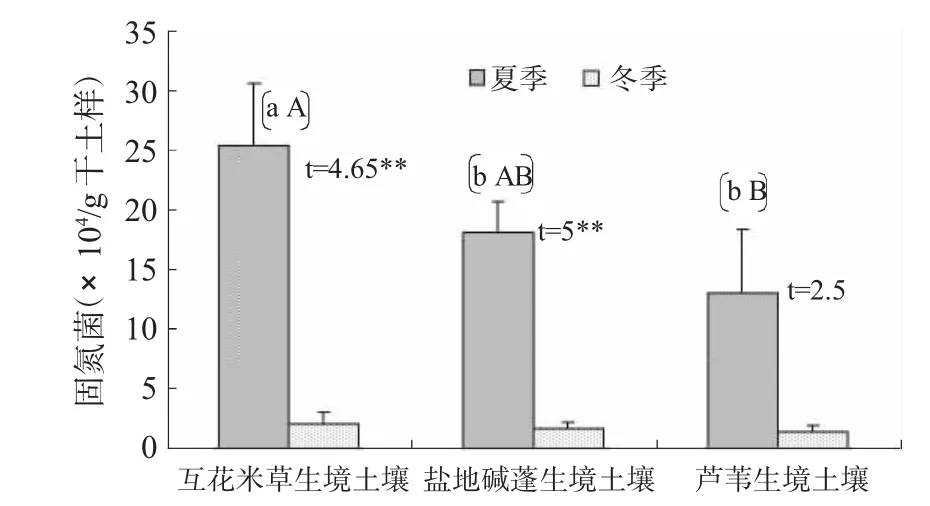
图5 不同盐生植物生境中土壤固氮菌数量差异
3 结论与讨论
3.1 结论
盐地碱蓬、芦苇及互花米草是江苏沿海滩涂中主要的盐生植物群落,它们生长在与农科院水稻生境明显不同的环境中,土壤性状的差异导致了土壤微生物数量的差异。农科院水稻田土壤熟化程度高,土壤营养丰富,因此土壤微生物数量较多,除真菌外,细菌、放线菌、纤维素分解菌和固氮菌等均明显高于盐生植物生境中的,在水稻田中,夏季的土壤中细菌数量、真菌数量和放线菌数量多于冬季的土壤,纤维素分解菌和固氮菌数量以冬季的土壤中大于夏季的。盐生植物生境主要特点是土壤盐分高,营养匮乏,海水浸润程度不一,从而导致不同盐生植物生境土壤微生物数量的差异。互花米草生境土壤盐分较高,海水浸润时间较长,土壤中除纤维素分解菌明显偏少外,其余菌种均多于盐地碱蓬生境土壤、芦苇生境土壤中的,且均以夏季的数量高于冬季的数量。盐地碱蓬主要生长在脱水的盐碱地中,土壤中除真菌数量略微高于其他两种植物生境土壤中的之外,其余菌种数量均居中间,少于互花米草生境土壤中的,高于芦苇生境土壤中的,也是夏季的数量多于冬季的数量。芦苇生境中盐分相对较低,土壤中除纤维素分解菌数量明显高于其他两种植物生境土壤中的之外,其余的数量均较少。
3.2 讨论
土壤是无机物、有机物和生物(主要是微生物)的有机复合体。土壤微生物作为土壤生态系统中的主要组成部分之一,是生态系统的分解者,是物质循环和能量交流的承担者,也是土壤中物质循环的动力,与土壤肥力、植物生长、土壤改良状况密切相关[10]。农科院稻田土壤经过多年改良,土壤有机质和氮源丰富,有机质含量高达0.419%,全氮含量0.21%,明显高于滩涂互花米草、碱蓬和芦苇生境土壤,稻田土壤微生物数量明显偏多,以细菌、纤维素分解菌和固氮菌的数量增加明显,其数量差异是与不同植物生长特性相适应的。
植物与土壤微生物之间通过植物的凋落物和植物根系分泌物建立起密切的联系,植物的多样性可以通过其凋落物和根系的分泌物导致植物和微生物之间的协同进化,促进土壤微生物的多样性。因此,土壤微生物与植物的相互作用主要表现在与植物根系和凋落物的相互作用。植物根系给土壤微生物提供了一个特殊的生态环境,某些土壤微生物可以通过与植物之间的种间关系影响植物发育、群落结构和演替[11]。滩涂芦苇生长环境土壤水分充足但少在流动水域,在秋冬季节常常茎枯叶落,给土壤微生物提供了丰富的营养源,尤其芦苇丰富的纤维素源,使得芦苇生境中纤维素分解菌数量明显较互花米草和碱蓬生境中的偏多。互花米草常常生长在流水环境,沿岸海水中含有丰富的生物,米草生境土壤中有机质含量较碱蓬、芦苇的高,细菌数量多少与土壤有机质含量大小趋势一致,此与高巍[12]、张恩平[13]和胡可[14]等研究结论一致。土壤盐分对微生物数量影响不明显,其可能对微生物种类产生影响。
土壤微生物数量有季节性差异,季节变化带来土壤微生物的生态环境变化,土壤微生物随季节的变化是明显的,季节对土壤微生物的影响是通过土壤温度、湿度、有机物的供应和植物生长状况等因素的综合作用形成的。滩涂米草、碱蓬和芦苇生境土壤中纤维素分解菌、放线菌数量季节性差异不明显,细菌、真菌和固氮菌的数量是夏季的明显大于冬季的,其影响因子主要是环境温度和营养源。
滩涂湿地土壤微生物动态是目前国内外研究热点之一,也是滩涂合理开发利用的基础,植物多样性影响土壤微生物多样性,从而进一步影响到土壤的理化性状,人类的生产与生活可以改变地面的植被组成和生长状况,植物群落的结构和组成的变化会导致植物物种组成的差异,并对土壤微生物产生重大影响,通过对滩涂主要盐生植物生境土壤微生物的研究,从而可以对植物群落演替过程的各环境因子的作用进行全面的研究。
[1] 裘江海.我国近代滩涂开发利用综述[J].水利发展研究,2006(3):26-29.
[2] 熊万英,王建.江苏沿海滩涂可持续发展研究[J].国土与自然资源研究,2004(4):52-54.
[3] 赵先丽,周广胜,周莉,等.盘锦芦苇湿地土壤微生物数量研究[J].土壤通报,2008,39(6):1376-1379.
[4] 康贻军,胡 健,董必慧,等.滩涂盐碱土壤微生物生态特征的研究[J].农业环境科学学报,2007,26(S):181-183.
[5] 叶温乐,何雪青,赵平芝,等.江苏盐城新洋港互花米草盐沼的微生物区系调查[J].中国农学通报,2007,23(8):420-424.
[6] 林学政,陈靠山,何培青,等.种植盐地碱蓬改良滨海盐渍土对土壤微生物区系的影响[J].生态学报,2006,26(3):801-807.
[7] 中国土壤学会农业化学专业委员会.土壤农业化学常规分析方法[M].北京:科学出版社,1983:12-60.
[8]中国科学院南京土壤研究所微生物室.土壤微生物研究法[M].北京:科学出版社,1985.
[9] 姚槐应,黄昌勇.土壤微生物生态学与实验技术[M].北京:科学出版社,2006.
[10] 樊盛菊,齐树亭,武洪庆,等.盐生植物根际对土壤中微生物数量和酶活性的影响[J].河北大学学报(自然科学版),2006,2(1):38-41.
[11] 李骁,王迎春.土壤微生物多样性与植物多样性[J].内蒙古大学学报(自然科学版),2006,37(6):708-713.
[12] 高巍,张淑红,张恩平,等.不同培肥方式对菜田土壤微生物生态特征的影响[J].沈阳农业大学学报,2009,40(2):140-143.
[13] 张恩平,高巍,张淑红,等.长期施肥条件下菜田土壤微生物特征变化[J].生态学杂志,2009,28(7):1288-1291.
[14] 胡可,李华兴,卢维盛,等.生物有机肥对土壤微生物活性的影响[J].中国生态农业学报,2010,18(2):303-306.
猜你喜欢
英语世界(2023年10期)2023-11-17 09:18:22
儿童故事画报·自然探秘(2022年3期)2022-04-27 00:17:10
齐鲁工业大学学报(2021年2期)2021-04-23 06:37:04
国际呼吸杂志(2019年22期)2019-12-09 09:20:36
水利科技与经济(2016年8期)2016-04-22 03:41:30
精品(2015年9期)2015-01-23 01:35:32
应用海洋学学报(2014年4期)2014-11-22 07:43:54
天然产物研究与开发(2014年6期)2014-04-27 14:15:56
中国医学科学院学报(2013年2期)2013-03-11 20:25:49
食品科学(2013年19期)2013-03-11 18:27:32
