
温度对等鞭金藻塔溪堤品系细胞密度和叶绿素荧光参数的影响
2011-03-20 03:42梁英范丽敏陈书秀王燕
大连海洋大学学报 2011年5期
梁英,范丽敏,陈书秀,王燕
(中国海洋大学海水养殖教育部重点实验室,山东青岛266003)
温度对等鞭金藻塔溪堤品系细胞密度和叶绿素荧光参数的影响
梁英,范丽敏,陈书秀,王燕
(中国海洋大学海水养殖教育部重点实验室,山东青岛266003)
以等鞭金藻塔溪堤品系Isochrysis galbana(Tahitian strain)为试验材料,采用Water-PAM叶绿素荧光仪测定了该藻一次性培养过程中,不同温度(5~40℃)对其细胞密度、叶绿素相对含量以及叶绿素荧光参数(PSⅡ最大光能转化效率Fv/Fm、光化学淬灭qP)的影响。结果表明:温度对等鞭金藻塔溪堤品系的叶绿素相对含量和细胞密度均有显著影响(P<0.05);在本试验条件下,等鞭金藻塔溪堤品系在5、10℃和40℃下不能生长繁殖,该藻生长的适温为15~35℃,最适温度为20~30℃,各处理组的相对生长率从高到低依次为25℃>20℃>30℃>35℃>15℃;温度对等鞭金藻塔溪堤品系各叶绿素荧光参数也有显著影响(P<0.05),从接种后第1天到试验结束,5、10℃和40℃处理组藻类的主要荧光参数(Fv/Fm、qP)均显著低于其它处理组(P<0.05),其中25℃和30℃处理组藻类的主要荧光参数总体上低于15℃、20℃和35℃处理组,但高于其它处理组;试验结束时将5、10℃和40℃处理组的藻细胞重新置于25℃下继续培养6 d,5℃和40℃处理组藻类的主要荧光参数不能恢复,而10℃处理组藻类的主要荧光参数可恢复正常。相关性分析结果表明:荧光参数、叶绿素相对含量以及细胞密度与温度的相关性随着培养时间的变化而变化,叶绿素相对含量与细胞密度呈显著的正相关关系(P<0.05)。
等鞭金藻塔溪堤品系;温度;细胞密度;叶绿素荧光参数
等鞭金藻具有个体小、无细胞壁、生长繁殖快、富含多不饱和脂肪酸等营养物质、易被消化吸收的特点,是水产经济动物的优质饵料[1-3]。目前育苗生产上常用的等鞭金藻有湛江等鞭金藻、等鞭金藻3011品系、等鞭金藻8701品系、等鞭金藻塔溪堤品系Isochrysis galbana(Tahitian strain)等,其中等鞭金藻塔溪堤品系最初是从塔溪堤(南太平洋的一个岛屿)养殖水体中分离筛选而得。虽然国内外一些学者对原种[4]及国内种质库保存的等鞭金藻塔溪堤品系[5]的适温范围进行了一些研究,但由于品系不同,研究结果差异较大。
研究表明,叶绿素荧光是光合作用的理想指标和探针。微藻体内的叶绿素荧光包含了丰富的与光合作用有关的生理信息,可以通过叶绿素荧光参数的变化来反映逆境对微藻光合作用某一过程的影响[6]。梁英等[7-8]采用叶绿素荧光技术对三角褐指藻和雨生红球藻的适温范围进行了研究,结果表明,可以通过某些叶绿素荧光参数的变化来确定这些微藻的适应温度,并可作为筛选耐高温或耐低温微藻的参考指标。到目前为止,关于不同温度对等鞭金藻塔溪堤品系叶绿素荧光参数影响方面的研究尚未见报道。本试验中,作者利用Water-PAM叶绿素荧光仪研究了在一次性培养等鞭金藻塔溪堤品系(MACC/H29)过程中,不同温度对其细胞密度、叶绿素相对含量和叶绿素荧光参数的影响,以及受胁迫后微藻在正常温度下荧光参数的恢复情况,旨在为该藻的培养及耐高(低)温品系的选育提供参考资料。
1 材料与方法
1.1 藻种及培养条件
等鞭金藻塔溪堤品系取自中国海洋大学微藻种质库。藻种的培养条件:采用f培养基,温度为
25℃,盐度为31,光照周期为16L∶8D,光照强度为100 μmol/(m2·s)。
1.2 方法
1.2.1 试验设计 试验在500 mL的三角烧瓶中进行,将处于指数生长期的藻种10 mL接种到新配制的f培养基中,培养液最终体积为400 mL,起始接种密度为10.92×104个/mL。试验中,温度分别设置为5、10、15、20、25、30、35、40℃,每个温度处理组设3个重复,其它培养条件与藻种培养条件相同。培养过程中不充气,每日随机调换三角烧瓶的位置并充分摇瓶5次,培养时间为10 d。
1.2.2 细胞密度的测定 每天定时取样,用血球计数板测定细胞密度。微藻相对生长率计算公式为
μ=(lnN-lnN0)/T,
其中:N0为T时间开始时的细胞数量;N为经过T时间后的细胞数量;T为藻类的生长时间(d)。每天用Water-PAM叶绿素荧光仪按照梁英等[7]的方法定时测叶绿素荧光参数(PSⅡ最大光能转化效率Fv/Fm、光化学淬灭qP)和叶绿素的相对含量。
1.2.3 藻类恢复培养试验 第1阶段温度试验结束后,将5、10℃和40℃处理组的微藻样品置于25℃下继续培养6 d,光照强度不变。每天定时取样,对叶绿素荧光的主要参数进行测定,以25℃处理组各荧光参数的最高值作为对照,观察藻类的恢复情况。
1.3 数据分析
用Sigmaplot 10.0软件作图。采用SPSS 11.5进行单因子方差分析,采用Duncan多范围检验法对处理组之间进行多重比较及相关性分析,以P<0.05表示差异显著。
2 结果
2.1 温度对等鞭金藻塔溪堤品系叶绿素荧光参数的影响
不同温度对等鞭金藻塔溪堤品系叶绿素荧光参数的影响见图1。单因子方差分析结果表明,温度对该藻的主要荧光参数Fv/Fm、qP均有显著影响(P<0.05)。多重比较结果表明,接种后第1天,5℃处理组的主要荧光参数基本下降为0;40℃处理组的主要荧光参数从接种后开始快速下降,第3天下降为0。10℃处理组的荧光参数Fv/Fm显著高于5℃和40℃处理组(P<0.05),但显著低于其它处理组;15℃处理组的荧光参数Fv/Fm在第6~10天显著高于其它处理组(P<0.05);25℃和30℃处理组的主要荧光参数总体上低于15℃、20℃和35℃处理组,未达到显著性差异,但显著高于其它处理组。qP的变化趋势均与温度有关,多数处理组的qP均呈现先下降后上升的趋势,5℃和40℃处理组的qP下降幅度最大,直接下降为0;20℃处理组的qP上升幅度最大。
等鞭金藻塔溪堤品系的荧光参数与温度之间的相关关系见表1。结果表明,荧光参数Fv/Fm、qP与温度的相关性和培养时间有关。温度为5~25℃时,第1~4天,Fv/Fm与温度呈极显著正相关(P<0.01);第1~6天时,qP与温度呈显著正相关(P<0.05)。25~40℃时,整个培养周期中, Fv/Fm、qP与温度均呈极显著负相关(P<0.01)。
等鞭金藻塔溪堤品系的荧光参数与细胞密度之间的相关关系见表2。从表2可见,荧光参数Fv/Fm、qP与细胞密度的相关性与培养时间有关,第2~7天,Fv/Fm与细胞密度呈显著正相关(P<0.05);第3~5天时,qP与细胞密度呈显著正相关(P<0.05)。
2.2 温度对等鞭金藻塔溪堤品系叶绿素相对含量和细胞密度的影响
温度对等鞭金藻塔溪堤品系叶绿素相对含量和细胞密度的影响见图1。单因子方差分析结果表明,不同温度对该藻的叶绿素相对含量和细胞密度均有显著影响(P<0.05)。多重比较结果表明:接种后第4~7天,25℃和30℃处理组藻类叶绿素的相对含量均显著高于其它处理组(P<0.05);接种后第7天,25℃处理组的细胞密度显著高于其它处理组(P<0.05);在整个培养周期中,5、10℃和40℃处理组藻类叶绿素的相对含量和细胞密度均显著低于其它处理组(P<0.05),不仅没有增加,反而有所减少,到第3天时,40℃处理组细胞密度下降为0,说明这3个温度下等鞭金藻塔溪堤品系不能生长繁殖;15℃处理组藻类叶绿素的相对含量和细胞密度均显著高于5、10℃和40℃处理组(P<0.05),但显著低于其它处理组(20、25、30、35℃),与接种时相比,培养结束时该处理组藻类叶绿素的相对含量和细胞密度增加幅度较少,分别为接种时的6.19和9.46倍。从图2可见,由于5、10℃和40℃处理组藻类没有生长繁殖,因此无法计算这3个处理组的相对生长率,其它各处理组相对生长率从高到低依次为25℃>
20℃>30℃>35℃>15℃,25℃处理组的相对生长率最高,但与20℃和30℃处理组之间差异不显著(P>0.05),与15℃、35℃处理组差异显著(P<0.05)。
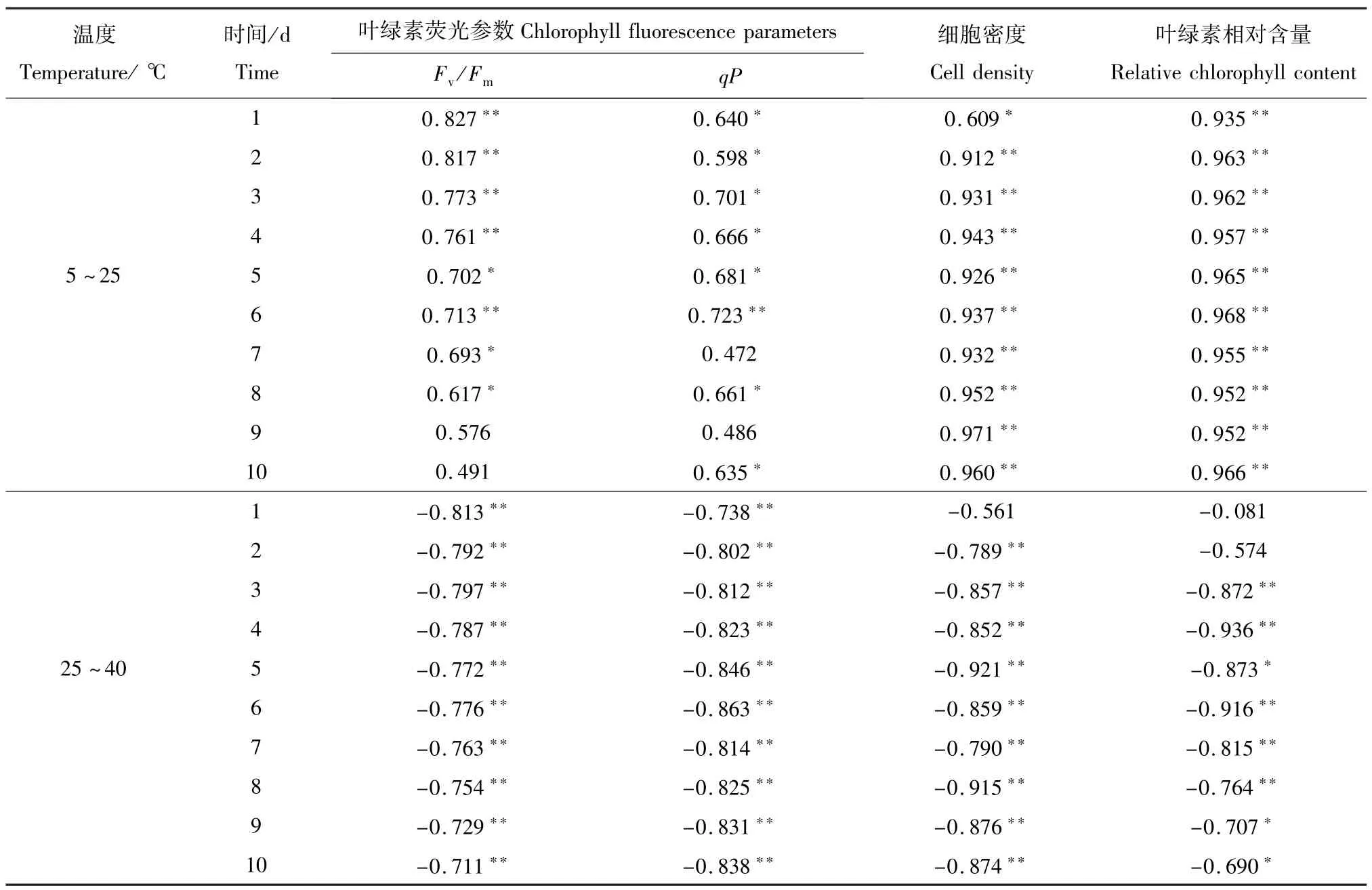
表1 等鞭金藻塔溪堤品系叶绿素荧光参数、细胞密度及叶绿素相对含量与温度的相关系数Tab.1 The correlation coefficients of chlorophyll fluorescence parameters,cell densities and relative chlorophyll content with temperature of Isochrysis galbana(Tahitian strain)
等鞭金藻塔溪堤品系叶绿素相对含量和细胞密度与温度间的相关关系见表1。结果表明,在5~25℃下,藻类叶绿素相对含量及细胞密度与温度呈显著的正相关关系(P<0.05或P<0.01),并随温度的增加而增加;25~40℃时,两者与温度的相关性与培养时间有关,第3~10天时,叶绿素相对含量及细胞密度与温度呈显著的负相关关系(P<0.05或P<0.01)。
从表2可见,整个培养周期中,该藻的叶绿素相对含量与细胞密度均呈显著的正相关关系。

表2 等鞭金藻塔溪堤品系叶绿素荧光参数及叶绿素相对含量与细胞密度的相关系数Tab.2 The correlation coefficients of chlorophyll fluorescence parameters and relative chlorophyll content with cell density of Isochrysis galbana(Tahitian strain)
2.3 藻类的恢复培养情况
将5、10℃和40℃处理组的藻类在温度为25℃、光照为100 μmol/(m2·s)的条件下恢复培养6 d后,5℃和40℃处理组的藻类完全不能恢复。从图3可见:10℃处理组藻类恢复培养3 d后,荧光参数(Fv/Fm、qP)分别比25℃处理组的最高值高出5.43%、5.89%。恢复培养6 d后,Fv/Fm恢复为25℃处理组最高值的96.87%,而qP却下降了6.27%,各参数基本恢复正常。
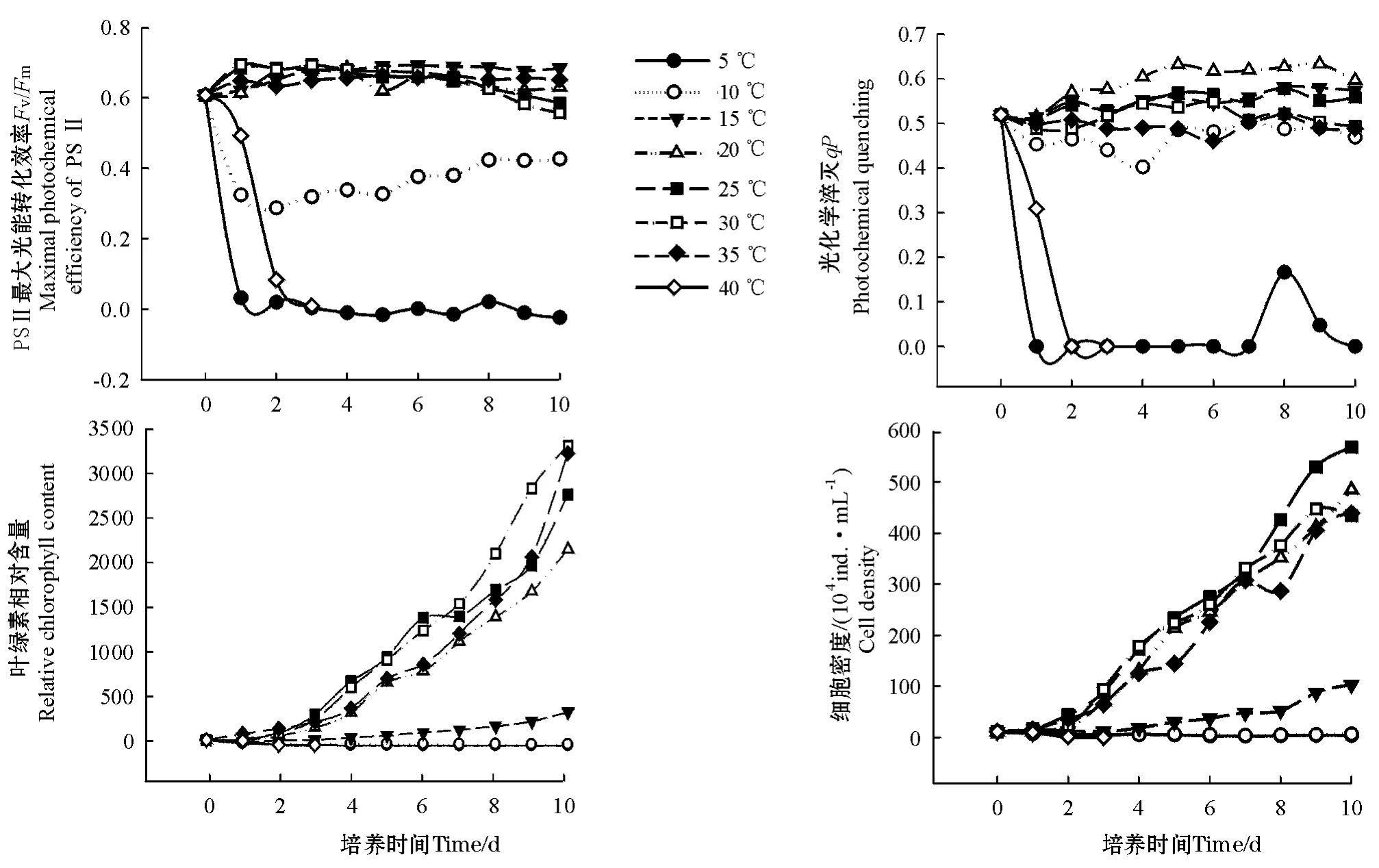
图1 不同温度对等鞭金藻塔溪堤品系叶绿素荧光参数(Fv/Fm,qP)、叶绿素相对含量及细胞密度的影响Fig.1 Effects of different temperatures on the chlorophyll fluorescence parameters,relative chlorophyll content and cell density Isochrysis galbana(Tahitian strain)

图2 不同温度下等鞭金藻塔溪堤品系的相对生长率Fig.2 Relative growth ratio of in the strain of Isochrysis galbana under different temperatures
3 讨论
本试验结果表明:等鞭金藻塔溪堤品系生长的适温为15~35℃,最适温度为20~30℃;在5、10℃和40℃下,该藻类的细胞密度和叶绿素相对含量不断减少,不能生长繁殖。这与Ukeles[9]认为原种等鞭金藻在低于8℃、高于27℃时不能繁殖的结果有所差异。据柳敏海等[5]报道,大地溪金藻在10~35℃均能生长和繁殖,最适温度为20~30℃,这与本试验中该藻在10℃下不能繁殖的结果不同。其原因可能是微藻品系不同而引起的,如Ukeles[9]用的是原种等鞭金藻,柳敏海等[5]用的是宁波大学生命学院藻种室保存的藻种,而本试验中用的是中国海洋大学微藻种质库保存的藻种。
叶绿素荧光技术是研究环境因子对植物光合作用影响的快速、灵敏、简捷的方法之一,荧光参数Fv/Fm、qP与逆境胁迫程度存在相关性。本试验结果表明,5℃和10℃低温胁迫对等鞭金藻塔溪堤品系叶绿素荧光参数影响较大,该温度下荧光参数均明显低于其他处理组,且5℃下该藻荧光参数下降为0。这表明低温胁迫下等鞭金藻塔溪堤品系PSⅡ最大光能转化效率下降,PSⅡ天线色素吸收的光能用于光化学电子传递的份额降低。低温对微藻光合作用的影响表现在许多方面。微藻受到低温胁迫时,反应中心进行代谢的途径中断,电子传递受阻,电子传递链中一部分电子与分子氧结合发生氧化还原反应,形成最初的反应氧形式——活性氧簇(ROS),ROS可能毁坏PSⅡ反应中心的蛋白复合物,引起光合作用的活性下降[10-11]。另外,低温
还会导致光抑制[12],引起类囊体膜流动性降低,对同化力的利用减少,因此,对脂质的双分子层和活性氧清除酶造成损伤,故影响了光合电子的传递效率和光合磷酸化过程[13]。植物吸收的光能除了用于光化学反应外,多余部分将耗散。耗散机制有多种形式,如热耗散机制、抗氧化机制(抗氧化剂和抗氧化酶)、PSⅡ反应中心失活和修复循环机制等。10℃处理组藻类的恢复培养试验结果表明,各荧光参数值能恢复到正常生长状态,说明该温度对等鞭金藻塔溪堤品系造成的是可逆性损伤,在合适的条件下可以恢复。因此,可以将等鞭金藻塔溪堤品系在10℃左右的低温下保存,需要时再放在合适的温度下进行活化,这样可以减少更换培养液的次数,避免因频繁更换培养液而引起的污染,并能保持种质遗传的稳定性,实现长期保存的目的[14-15]。

图3 10℃下培养10 d后等鞭金藻塔溪堤品系恢复培养后的叶绿素荧光参数(温度为25℃,光强为100 μmol/(m2·s))Fig.3 Chlorophyll fluorescence parameters of Isochrysis galbana(Tahitian strain)during recovery(25℃,100 μmol/(m2·s))after cultured under 10℃for ten days.
本试验结果表明,高温处理组藻类(35℃)接种后2~5 d,其荧光参数Fv/Fm、qP显著高于5℃、10℃处理组,但低于15、20、25、30℃处理组,说明35℃处理组其PSⅡ的最大光能转化效率和光化学淬灭都受到了抑制,但是其抑制程度小于10℃处理组;40℃处理组藻类接种后3 d,其荧光参数Fv/Fm、qP均下降为0,且无法恢复,说明该温度对等鞭金藻塔溪堤品系造成的是不可逆性损伤,40℃下该藻根本无法生长繁殖。光合作用是微藻对高温最敏感的生理过程之一。高温主要破坏PSⅡ,并对PSⅡ的反应中心产生影响,可引起藻类捕光色素复合体同PSⅡ的反应中心分离,导致类囊体膜解垛叠[16],使捕光色素复合体的结构发生改变[17]、放氧复合体失去活性[18]以及Q→QB的电子传递被抑制等。
本试验结果还表明,随着培养时间的增加,35℃处理组叶绿素相对含量和细胞密度不断增加,到培养结束时,分别增加到接种时的53.8和40.2倍,说明该藻在35℃条件下可以较好地生长繁殖,属于耐高温品系。这种特性正好符合南方及北方夏季对耐高温金藻品系的要求。但40℃处理组藻类叶绿素的相对含量和细胞密度在培养3 d后几乎下降为0,且无法恢复。因此在本试验条件下,该藻种耐高温的极限为35~40℃。虽然该藻可以耐较高温度,但南方夏季水产养殖生产中还需要耐更高温度的金藻品系,因此,为满足南方水产养殖生产的需要,还需要筛选耐更高温度的金藻品系。叶绿素荧光技术是一种快速、灵敏、对细胞无伤害的检测技术,利用该技术在筛选麦类作物耐盐品种方面已显示出较好的应用前景,可以用于盐胁迫下叶绿素荧光参数的变化,以此来筛选小麦和大麦的耐盐基因型[19-20]。本试验中,等鞭金藻塔溪堤品系的荧光参数Fv/Fm以及叶绿素相对含量与细胞密度的相关性较好,可以用来评价该藻对温度的适应性。目前,仅对少量的微藻品种进行了这方面的研究。随着人们对各种环境因子(温度、盐度等)胁迫下叶绿素荧光参数变化机理的进一步了解,以及荧光参数测定技术的进一步完善,活体叶绿素荧光测定技术将在微藻耐性生理研究以及微藻育种中发挥越来越重要的作用。
[1] Cañavate J P,Lubińn L M.Some aspects on the cryopreservation of microalgae used as food for marine species[J].Aquaculture,1995, 136:277-290.
[2] Fernández E M,Salmón H A,Dávalos C R.Ingestion and digestion of 10 species of microalgae by winged pearl oyster Pteria sterna (Gould,1851)larvae[J].Aquaculture,2004,230:417-423.
[3] Wikfors G H,Patterson G W.Differences in strains of Isochrysis of importance to mariculture[J].Aquaculture,1994,123:127-135.
[4] Kain J M,Fogg G E.Studies on the growth of marine phytoplankton
[J].Mar Biol,1958,37:781-788.
[5] 柳敏海,张春丹.大溪地金藻培养的生态条件研究[J].水产科学,2005,24(3):10-12.
[6] 冯建灿,胡秀丽,王训申.叶绿素荧光动力学在研究植物逆境生理中的应用[J].经济林研究,2002,20(4):14-18.
[7] 梁英,陈书秀,田传远,等.温度对三角褐指藻叶绿素荧光特性及生长的影响[J].中国海洋大学学报,2008,38(3):377-383.
[8] 梁英,陈书秀.温度对雨生红球藻叶绿素荧光特性及虾青素含量的影响[J].海洋湖沼通报,2009(3):112-120.
[9] Ukeles R.The effect of temperature on the growth and survival of several marine algal species[J].Biol Pharm Bull,1961,120:255-263.
[10] 张守仁.叶绿素荧光动力学参数的意义及讨论[J].植物学通报,1999,16(4):444-448.
[11] Clare D C,Rabinowitch H D,Fridovich I.Superoxide dismutase and chilling injury in Chlorella ellipsoidea[J].Arch Physiol Biochem,1984,231(1):158-163.
[12] Papageogious G.Chlorophyll fluorescence an intrinsic probe of photosynthesis[M]//Biogetics of photosynthesi.New York:Academic press,1975:319-371.
[13] Mallick N,Mohn F H.Use of chlorophyll fluorescence in metal stress research:a case study with the green microalgae Scenedesmus[J].Ecotoxicol Environ Saf,2003,55:64-69.
[14] 张跃群,石斌,陆德祥,等.3种优质海洋微藻的低温保存研究[J].江苏农业科学,2009(6):314-317.
[15] 王培磊,宫庆礼,麦康森,等.两种海洋单胞藻浓缩与保存效果的研究[J].海洋湖沼通报,2001(4):12-19.
[16] 黄仿,武宝玕.热胁迫对球等鞭金藻作用机制的叶绿素荧光研究[J].广西师范大学学报,1995,11(2):72-76.
[17] 武宝轩,韩志国,藏汝波.高温胁迫对海洋红藻及绿藻叶绿素荧光的影响[J].暨南大学学报,2002,23(1):108-112.
[18] Srivastava A,Guisse B,Greppin H,et al.Regulation of antenna structure and electron transport in photosystem II of Pisumsatium under elevated temperature probed by the fast polyphasic chlorophyll a fluorescence transient:OK JIP[J].Biochim Biophys Acta, 1997,1320:95-106.
[19] Laing L,Utting S D.The influence of salinity on the production of two commercially important unicellular marine algae[J].Aquaculture,1980,21:79-86.
[20] Belkhodja R,Morales F,Abadía A,et al.Effects of salinity on chlorophyll fluorescence and photosynthesis of barley(Hordeum vulgare L.)grown under a triple-line-source sprinkler system in the field[J].Photosynthetica,1999,36:375-387.
Effects of temperature on cell density and chlorophyll fluorescence parameters in Tahitian strain of alga Isochrysis galbana
LIANG Ying,FAN Li-min,CHEN Shu-xiu,WANG Yan
(The Key Laboratory of Mariculture,Ministry of Education,Ocean University of China,Qingdao 266003,China)
The effects of temperatures(5-40℃)on cell density,relative chlorophyll content and chlorophyll fluorescence parameters(maximal effective quantum yield of PSⅡin the dark,Fv/Fm;the photochemical quenching coefficient qP)were examined in Tahitian strain of Isochrysis galbana in batch cultures by Water-PAM chlorophyll fluorometer.One-way analysis of variance showed that temperature had significant effects on the cell density and relative chlorophyll content of the alga(Tahitian)during the experiment(P<0.05).Under this experimental condition,the proper growth temperature for the alga was 15-35℃,the optimal growth temperature of 20-30℃.The alga was found to cease growing and developing under 5,10℃and 40℃.The relative growth ratio under different temperatures was arranged as 25℃>20℃>30℃>35℃>15℃.Temperature also had significant effects on the fluorescence parameters of the alga.The main parameters(Fv/Fm,qP)were significantly lower under 5,10℃and 40℃than those in other treatments from day 1 to the end of this experiment.At 25℃and 30℃,however,the above main fluorescence parameters were generally lower than those at 15℃,20℃and 35℃,but higher than others treatments.After cultured at 5,10℃and 40℃for ten days,the microalga was re-cultured in 25℃,100 μmol/(m2·s)conditions for 6 days,all the parameters in 5℃and 40℃treatment were not recovered absolutely, while those in 10℃treatment were recovered to the normal level.The cell density,relative chlorophyll content and fluorescence parameters exhibited some correlations with temperatures,while the pattern and strength were varied with the temperature range and the culture period.Moreover,significant positive correlations between chlorophyll content and cell density were found during the experiment.
Isochrysis galbana;temperature;cell density;chlorophyll fluorescence parameter
Q945.78
A
2095-1388(2011)05-0422-06
2010-12-10
“十一五”国家科技支撑计划项目(2006BAD09A03)
梁英(1967-),女,博士,教授。E-mail:yliang@ouc.edu.cn
猜你喜欢
当代水产(2021年8期)2021-11-04
军事文摘(2020年20期)2020-11-16
阅读(科学探秘)(2020年8期)2020-11-06
园林科技(2020年2期)2020-01-18
山东林业科技(2019年2期)2019-06-03
中国果业信息(2019年1期)2019-01-05
今日农业(2019年10期)2019-01-04
生物学教学(2017年9期)2017-08-20
环境科技(2016年2期)2016-11-08
广西林业科学(2016年2期)2016-03-20
