
保加利亚乳杆菌H+-ATPase缺陷型菌株延缓后酸化及发酵性质的研究
2011-03-07 04:55庞启亮霍贵成
东北农业大学学报 2011年2期
庞启亮,霍贵成
(东北农业大学乳品科学教育部重点实验室,哈尔滨 150030)
发酵酸奶由于具有丰富的营养价值和良好的保健功能,普遍被消费者认可,其产销量迅猛增长。但在产品贮存、运输、销售、食用前这一过程中,乳酸菌仍能分解乳糖,产生乳酸,发生后酸化(Postacidification)。酸奶储存时的后酸化作用使酸奶的风味变化,pH降低,甚至达到消费者难以接受的程度。防止酸奶后酸化的方法多种多样,如:调节球杆看比例,增加球菌比例;添加Nisin,抑制保加利亚乳杆菌的代谢。然而在乳酸菌中,控制后酸化最好的方法是控制保加利亚乳杆菌的生长和能量代谢[1]。ATPase是控制保加利亚乳杆菌能量代谢的一个关键酶[2],H+-ATPase维持细胞内pH稳定的产生H+离子梯度,并驱动细胞糖和氨基酸的运输[3]。Kanner等利用新霉素选择不同ATPase活性的乳酸菌,原理在于新霉素进入细胞,需要H+-ATPase的协助,但细胞内的新霉素对细胞有抑制作用,低活性H+-ATPase运输新霉素的能力弱,对新霉素富集作用低,在筛选培养基中能够生长,而正常菌株H+-ATPase运输相对较多新霉素,抑制细胞生长[4]。
本试验利用新霉素[5]筛选德氏乳杆菌保加利亚亚种(Lactobacillus delbrueckii subsp.bulgaricus)H+-ATPase是原有亲本活性的60%[6],由于H+-ATPase是细胞代谢关键的酶,所以筛选菌株相对较弱后酸化能力的同时生长性能降低,通过高密度发酵,筛选适宜培养基并提高筛选菌种生物量得率成为发酵剂开发的另一重点。通过对筛选菌株后酸化活性检测,并筛选出适宜增殖培养基,为保加利亚乳杆菌H+-ATPase酶缺陷型菌株延缓后酸化发酵剂的开发奠定基础。
1 材料与方法
1.1 材料
菌种:德氏乳杆菌保加利亚亚种(Lactobacillus delbrueckii subsp.bulgaricus)L15,KLDS 1.9201,由东北农业大学乳品重点实验室提供;德氏乳杆菌保加利亚亚种L15-4,KLDS 1.9201-4,由东北农业大学乳品科学教育部重点实验室利用新霉素筛选自KLDS 1.9201;德氏乳杆菌保加利亚亚种LW-1自市售酸奶中分离(对照);嗜热链球菌(Streptococcus.thermophilus)S18,KLDS 3.9210由东北农业大学乳品科学教育部重点实验室提供。
脱脂乳配制:脱脂乳溶于蒸馏水12.6%W/V,90℃,10 min番茄汁[7]:市售,无病虫害及机械损伤。新鲜番茄→清洗→热烫(90~95℃,3 min)→榨汁→调配→灭菌备用。
MRS液体及MRS固体培养基,分别用于菌种活化及活菌计数[8]。
1.2 仪器
梅特勒-托利多Delta 320 pH计、超低温冰箱(-80℃)、GL-21M高速冷冻离心机、BCN1360型生物洁净工作台、SPX-150B生化培养箱、3.7 L瑞士比欧发酵罐。
1.3 方法
1.3.1 菌种和培养条件
德氏乳杆菌保加利亚亚种L15,德氏乳杆菌保加利亚亚种L15-4,德氏乳杆菌保加利亚亚种LW-1,MRS培养基中37℃培养18 h,4℃低温离心5000 r·min-1,10 min,生理盐水洗涤两次,灭菌脱脂乳悬浮,-80℃储存。S18在M17培养基中37℃培养18 h,4℃ 低温离心5000 r·min-1,10 min,生理盐水洗涤2次,灭菌脱脂乳悬浮,-80℃储存。
1.3.2 发酵酸奶
德氏乳杆菌保加利亚亚种L15,德氏乳杆菌保加利亚亚种L15-4,德氏乳杆菌保加利亚亚种LW-1分别接种5.0×106cfu·mL-1,42℃发酵至pH 4.8左右,分别在 4、20 °C 保存,0、3、6、9、12、15、18、21 d检测pH和滴定酸度。
L15与 S18分别接种5.0×106cfu·mL-1,德氏乳杆菌保加利亚亚种L15-4与S18分别接种5.0×106cfu·mL-1,德氏乳杆菌保加利亚亚种LW-1与S18分别接种5.0×106cfu·mL-1,42 ℃发酵至pH 4.8左右,分别在4、20℃保存,0、3、6、9、12、15、18、21 d检测pH和滴定酸度。
1.3.3 高密度发酵
以MRS培养基为基础培养基,37℃,5%接种量,脱脂乳、酵母粉、番茄汁3因素3水平,正交设计L9(33),方案见表1。
2 结果与分析
2.1 单一菌株发酵酸奶4℃储存的后酸化
结果见图1、2。
由图1、2可知,单一杆菌5.0×106cfu·mL-1接种量条件下,4°C储存L15后酸化最严重,12 d后pH降至4.16;21 d,pH降至4.10,滴定酸度增加340T;LW-1在4℃储存21 d,pH降至4.29,滴定酸度增加240T;筛选菌株L15-44℃储存21 d,pH降至4.40,滴定酸度增加200T。L15-4由于H+-ATPase活性弱,储存时产酸能力降低,远低于亲本L15,这是由于H+-ATPase参与维持细胞内正常pH,H+-ATPase活性弱,外界H+浓度对L15-4影响更显著,细胞代谢受阻,产酸能力降低,同时验证L15与对照组LW-1产酸能力差别,L15产酸能力更强,后酸化严重;筛选菌株L15-4后酸化能力弱于照组LW-1。
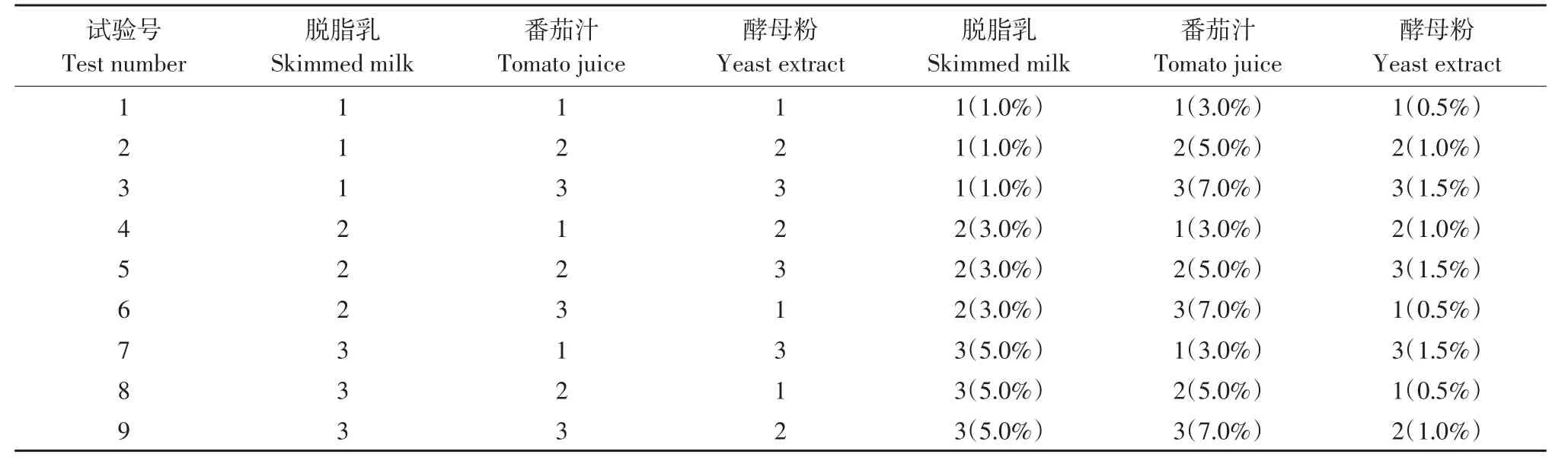
表1 增殖培养L9(33)试验方案Table1 Media of the multiplication to L9(33)
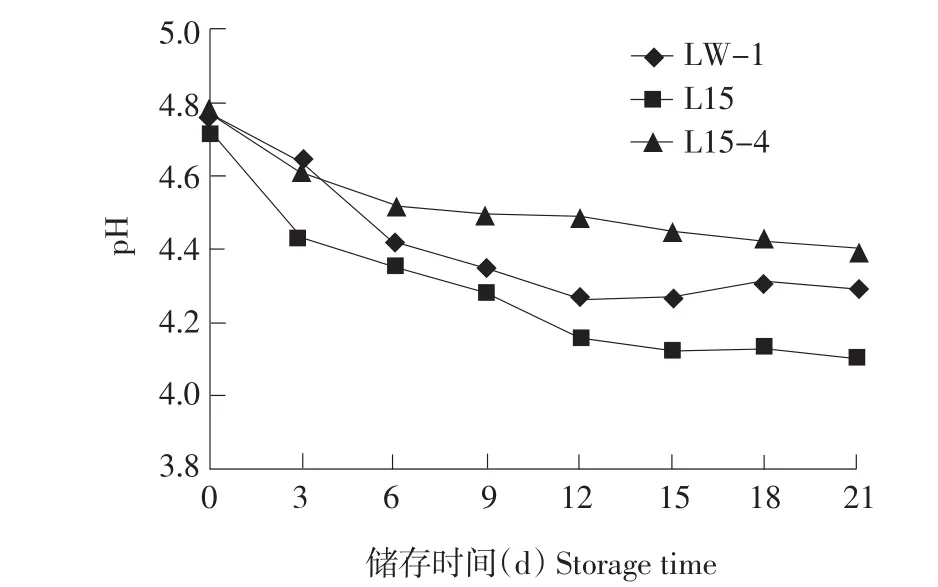
图1 单一发酵剂4℃储存pH变化Fig.1 pH variation of single starter fermented yogurt stored at 4℃
2.2 单一菌株发酵酸奶20℃储存的后酸化
酸奶在有相对完整制冷设备条件下,储存过程中,储存温度在4~10℃之间[10],检测在不完全具备制冷条件下,20℃储存后酸化状况,如图3、4所示,20℃储存L15后酸化比4℃更严重,21 d pH降至3.89,滴定酸度增加500T;LW-120℃储存21 d pH降至4.06,滴定酸度增加390T;筛选菌株L15-4在20℃储存21 d pH降至4.30,滴定酸度增加240T。虽然在20℃储存条件下L15-4弱后酸化能力依然明显,但L15-4在制作酸奶过程中,凝乳时间5 h,远高于亲本及对照菌株LW-1。
2.3 混合菌株发酵酸奶4℃储存的后酸化

图2 单一发酵剂4℃储存滴定酸度变化Fig.2 Titration acidity variation of single starter fermented yogurt stored at 4℃
混合球菌,分别接种5.0×106cfu·mL-1,在4℃储存,如图5、6所示:L15+S18混合菌株,21d储存pH降至4.01,滴定酸度增加440T;LW-1+S18混合菌株21 d储存pH降至4.14,滴定酸度增加310T;L15-4+S18混合菌株21 d储存pH降至4.26,滴定酸度增加230T。酸奶是由保加利亚乳杆菌与嗜热链球菌混合发酵而来,二者彼此相互促进生长,混合菌株发酵酸奶与单一保加利亚乳杆菌发酵酸奶性质不完全一致,然而混合菌株发酵,筛选菌株L15-4后酸化程度较亲本低。
2.4 混合菌株发酵酸奶20℃储存的后酸化
20℃储存,混合菌株发酵,如图7、8所示:L15+S18混合菌株,21 d储存pH降至3.87,滴定酸度增加640T;LW-1+S18混合菌株21 d储存pH降至4.02,滴定酸度增加410T;L15-4+S18混合菌株21 d储存pH降至4.16,滴定酸度增加300T。模拟现实酸奶储存条件,筛选菌株L15-4弱后酸化能力依然明显。
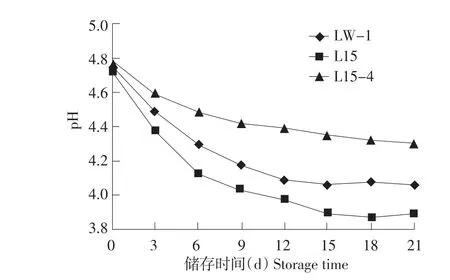
图3 单一发酵剂20℃储存pH变化Fig.3 pH variation of single starter fermented yogurt stored at 20℃
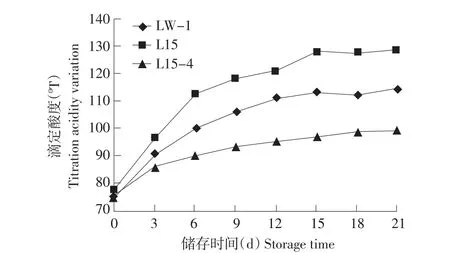
图4 单一发酵剂20℃储存滴定酸度变化Fig.4 Titration acidity variation of single starter fermented yogurt stored at 20℃
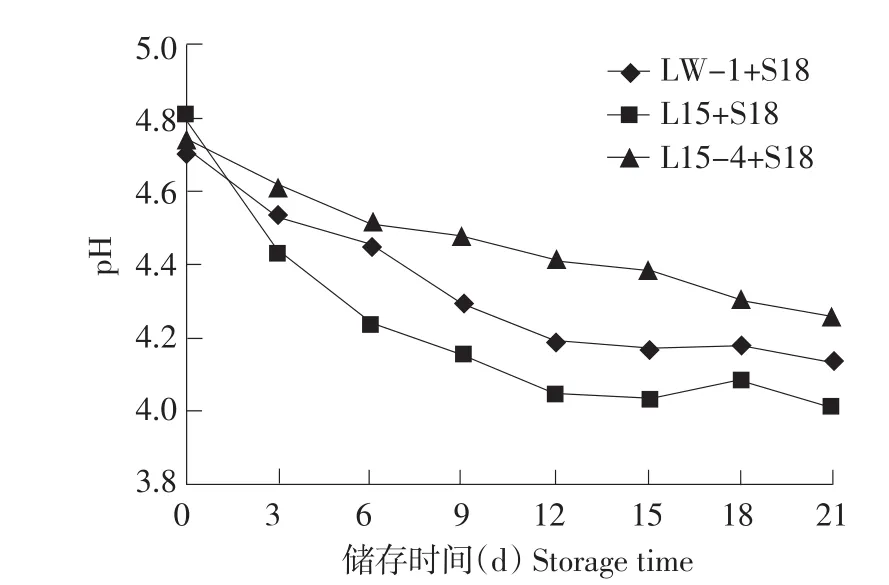
图5 混合发酵剂4℃储存pH变化Fig.5 pH variation of fermentation with stored at 4℃

图6 混合发酵剂4℃储存滴定酸度变化Fig.6 Titration acidity variation of fermentation with stored at 4℃
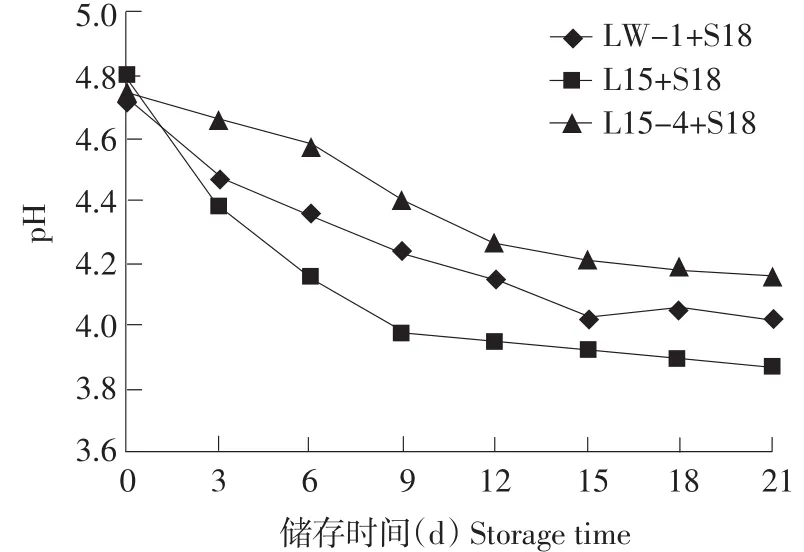
图7 混合发酵剂20℃储存pH变化Fig.7 pH variation of fermentation with stored at 20℃
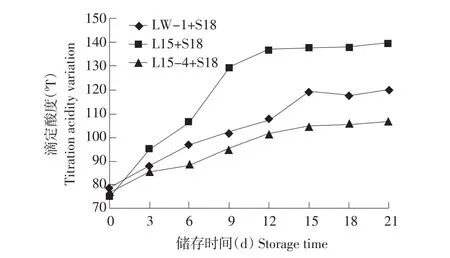
图8 混合发酵剂20℃储存滴定酸度变化Fig.8 Titration acidity variation of fermentation with stored at 20℃
2.5 正交试验设计筛选最适培养基
如表2所示,极差Rj的意义:Ki:i指某一特定因子第i个水平极差,Ki等于该因子第i个水平试验结果之和,Rj=K(最大值)-K(最小值),Rj越大,说明该因素的水平变化对试验指标的影响越大,该因素越重要;反之,Rj越小,该因素越不重要。由此可以根据Rj的大小顺序排出因素的主次,番茄汁是影响菌体高密度发酵的主要因素,脱脂乳次之,酵母粉对高密度发酵的影响最小。用正交表安排试验[12],最佳培养基组合为5号:脱脂乳3%,番茄汁5%,酵母粉0.5%,最高菌密度1.44×109cfu·mL-1。
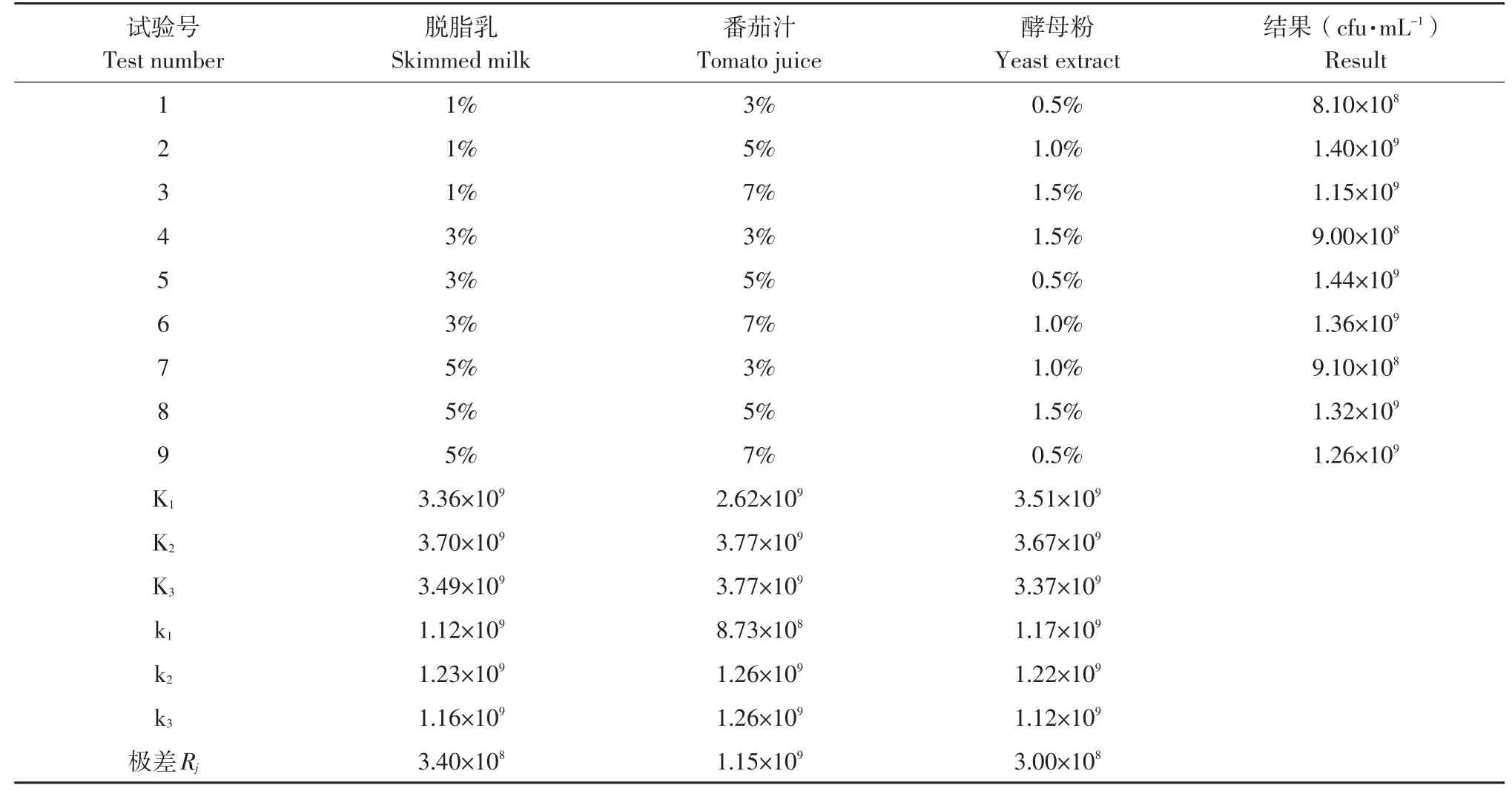
表2 增殖培养L9(33)试验结果Table2 Results of the multiplication to L9(33)
3 讨论与结论
L15-4 H+-ATPase的活力比亲本L15降低了60%,通过对新霉素筛选保加利亚乳杆菌菌株发酵酸奶,与亲本、对照组比较,后酸化明显减弱。H+-ATPase维持乳酸菌正常细胞内pH和物质运输,是乳酸菌正常生长的重要酶[2],并且保加利亚乳杆菌葡萄糖、谷氨酸盐、天冬氨酸盐等运输依赖H+-ATPase协助运输,H+-ATPase活性较弱,维持细胞内外H+浓度差的能力降低,细胞外H+浓度变化对H+-ATPase型保加利亚乳杆菌抑制作用更明显,并且小分子物质运输能力降低,细胞正常代谢受到影响,所以保加利亚乳杆菌对环境条件变化更敏感,存储时,不适宜pH条件,保加利亚乳杆菌对此反应更强烈,产酸能力降低,所以H+-ATPase型保加利亚乳杆菌后酸化能力降低。同时L15-4发酵酸奶过程中产酸性能、高密度发酵过程中生长性能受到H+-ATPase的活力的影响,相应地降低。H+-ATPase弱化保加利亚乳杆菌高密度发酵时,最高菌密度达1.44×109cfu·mL-1,低于其亲本2.15×109cfu·mL-1[13]。虽然L15-4 H+-ATPase活性降低带来诸如:凝乳时间延长,生产成本增加等不利因素,作为低档酸奶发酵剂不具有优势,然而在益生菌产品中具有广阔的应用前景,益生菌产品市场将成为酸奶市场的主流产品,由于益生菌中双歧杆菌的存活率受pH影响非常大[11],H+-ATPase型保加利亚乳杆菌后酸化能力弱化,相应提高益生菌产品中益生菌存活率,H+-ATPase型保加利亚乳杆菌生产成本高,凝乳时间长,培养条件苛刻等不利条件的影响依然低于益生菌存活率提高带来的收益。
[1]Mollet B.Genetically improved starter strains:opportunities for the dairy industry[J].International Dairy Jounal,1999(9):11-15.
[2]Kuban B J,Nilsson D,Kuipers O P,et al.The membrane-bound H+-ATPase complex is essential for growth of Lactococcus lactis[J].Journal of Bacteriology,2000,182(17):4738-4743.
[3]Kobayashi H.A proton-translocating ATPase regulates pH of the bacterial cytoplasm[J].Journal of Biological Chemistry,1985,260:72-76.
[4]Kanner B I,Gutnick D L.Use of neomycin in the isolation of mutants blocked energy conservation in Escherichia coli[J].Journal of Bacteriology,1971,111:287-289.
[5]Rosen B P.Restoration of active transport in an Mg2+-adenosin-etriphosphatase-deficient mutant of Escherichia coli[J].Journal of Bacteriology,1973,116:1124-1129.
[6]Sakamoto K,Veen H W,Saito H,et al.Membrane-bound ATPase contributes to hop resistance of Lactobacillus brevis[J].Applied and Environmental Microbiology,2002,68:5374-378.
[7]Blank L M,Koebmann B J,Michelsen O,et al.Hemin reconstitutes proton extrusion in an H+-ATPase-negative mutant of Lactococcus lactis[J].American Society for Microbiology,2001,183(22):670-677.
[8]刘飞,杜鹏,王玉堂,等.保加利亚乳杆菌H+-ATPase缺陷型菌株的筛选[J].微生物学报,2009,49(1):38-43.
[9]杨洁彬,凌代文,郭兴华,等.乳酸菌-生物学基础及应用[M].北京:中国轻工业出版社,1999:139-176,187-202.
[10]国际乳品联合标准IDF 117:1983.酸奶特征微生物的菌落计数[S].北京:中国乳品工业,1990,18(4):179-182.
[11]Ongol M P,Sawatari Y,Ebina Y,et al.Yoghurt fermented by Lactobacillus delbrueckii subsp.bulgaricus H+-ATPase-defective mutants exhibits enhanced viability of Bifidobacterium breve during storage[J].International Journal of Microbiology,2007,116:358-366.
[12]张勤,张启能.生物统计学[M].北京:中国农业大学出版社,2002:220-223.
[13]李艾黎.酸奶菌种发酵工艺研究及优质乳品发酵剂的研制[D].哈尔滨:东北农业大学,2006:76-77.
猜你喜欢
中国土壤与肥料(2021年5期)2021-12-02
云南化工(2020年11期)2021-01-14
河北地质(2017年2期)2017-08-16
中国乳品工业(2015年12期)2015-12-16
中国酿造(2014年9期)2014-03-11
食品工业科技(2014年23期)2014-03-11
中国乳业(2013年10期)2013-08-31
俄罗斯问题研究(2012年1期)2012-03-25
俄罗斯问题研究(2012年1期)2012-03-25
俄罗斯问题研究(2012年1期)2012-03-25
